Типология ареалов
Типология ареалов
Каждый вид имеет свой собственный ареал, и в природе практически невозможно найти два абсолютно одинаковых по площади, расположению или по форме ареала, за исключением мелких островов, где различные виды животных могут населить всю его территорию, и тогда их ареалы будут изображаться на карте одинаково. Тем не менее все разнообразие ареалов можно свести к какому-то ограниченному числу типов.
Для номенклатуры ареалов важно использовать 3 оси координат, или 3 составляющие ареала: широтную (распространение с севера на юг), долготную (с запада на восток) и высотную (вертикальную, сверху вниз).
Широтная составляющая ареала (зональное распространение) является его основным географическим признаком. Она определяется в первую очередь климатическими, особенно температурными, факторами, прямо или косвенно влияющими на распространение большинства видов. Границы ареалов таких видов на определенном протяжении совпадают с границами ландшафтной зоны, хотя зональная приуроченность каждого из них может значительно меняться на всем протяжении ареала.
Высотная составляющая ареала (вертикальная поясность) в определенной степени аналогична широтной, поскольку также определяется главным образом температурными градиентами. По характеру вертикального распространения можно различить альпийские и монтанные (горные) виды. Последние заселяют среднегорья. Выделяют и промежуточные между ними — субальпийские формы. Часто приходится сталкиваться с равнинно-горным расселением. Примером может служить арктоальпийское или бореомонтанное распространение видов (рис. 116).
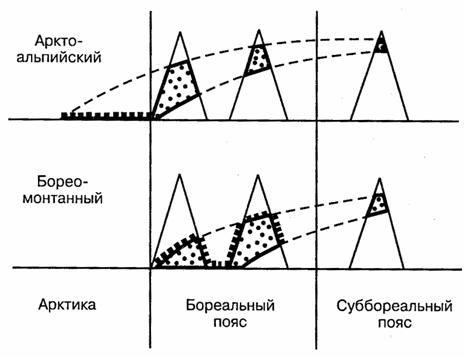
Рис. 116. Основные типы равнинно-горного
распространения животных (по К.Б.Городкову, 1984)
Долготная составляющая ареала определяется степенью удаленности от океана, обусловливающей характер климата. Границы ареалов могут быть связаны с континентальным или морским, аридным или гумидным климатом. В пределах гумидной части Евразии, к примеру, наиболее существенны следующие границы:
между Западной и Восточной Европой, по Уралу, по западному краю Среднесибирского плоскогорья («линия Иогансена»), в районе Байкала и по водоразделу бассейна Охотского моря.
Комбинация составляющих ареала позволяет охарактеризовать тип распространения и создать описательные классификации ареалов.
В основу классификаций должно быть положено одно из основных свойств ареалов — их размеры. Можно построить иерархическую систему ареалов — от узколокальных до глобальных (рис. 117). Наряду с очень узкими, занимающими территории в несколько квадратных километров и меньше, встречаются широкие ареалы, которые охватывают целые регионы, материки и даже весь земной шар. Как правило, наземные животные чаще имеют узкие ареалы, морские же — широкие. Иногда вид ограничен в своем распространении локальным районом, естественной географической (или фаунистической) областью. Такие виды называются эндемичными для данной области или просто эндемиками.
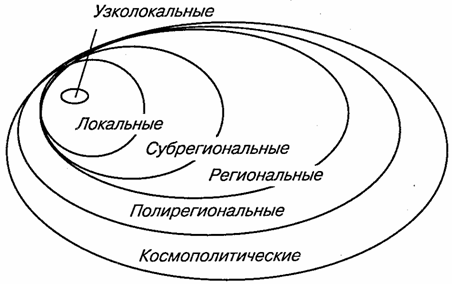
Рис. 117. Схема пространственного совмещения ареалов
различных размерных групп (по К. Б. Городкову, 1983)
Особенно узкие ареалы типичны для видов, распространение которых ограничено какими-нибудь непреодолимыми преградами. Это прежде всего островные либо пещерные формы, обитатели горных долин или, наоборот, верхних зон горных хребтов. Нередки весьма узкие ареалы у нелетающих видов насекомых. Например, жужелицы-брызгуны обитают на Кавказе в пределах одного-двух хребтов: Carabus polychrous населяют Бзыбский хребет в Абхазии, С. komarovi — Сванетию, С. cordicollis встречаются только на Эльбрусе. Аналогичные ареалы у бескрылых жуков-листоедов рода Oreomela в горах Тянь-Шаня и Памиро-Алтая. Так, О. medvedevi живет только в одном районе Шугнанского хребта на Памире, О. transalaica — на Заалайском хребте, О. bergi — в альпийской зоне Туркестанского хребта в верховьях реки Исфары. Все без исключения виды этого рода, а их больше 50, имеют очень узкие эндемичные ареалы. Исключительно узкие ареалы у трогло-бионтов, постоянно живущих в пещерах, причем это касается как наземных, так и пресноводных видов. Итак, все узкие ареалы свойственны эндемикам.
Локальные ареалы, постепенно расширяясь, переходят в субрегиональные и региональные. Если вид распространен на всей площади региона, то его ареал называют трансрегиональным (рис. 118).
Еще более широкие ареалы, так называемые полиили мультирегиональные, включают в себя несколько (не менее 2) фаунистических областей, часто располагающихся на разных материках (рис. 119, 120). Наконец, различают космополитные, или всесветные, ареалы (рис. 121). Они характерны чаще всего для морских животных, таких как кашалот (Physeter), дельфин (Grampus gri-seus), касатка (Orcinus orca). Из наземных животных космополитами стали определенные насекомые-синантропы. С некоторой натяжкой к космополитам можно отнести сокола-сапсана.
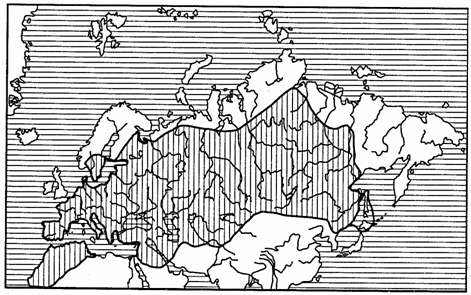
Рис. 118. Трансрегиональный ареал бабочки-боярышницы
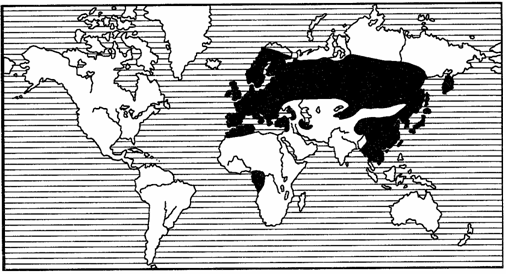
Рис. 119. Полирегиональный ареал пластинчатоусых жуков
рода Trichius (из С.И.Медведева, 1960)
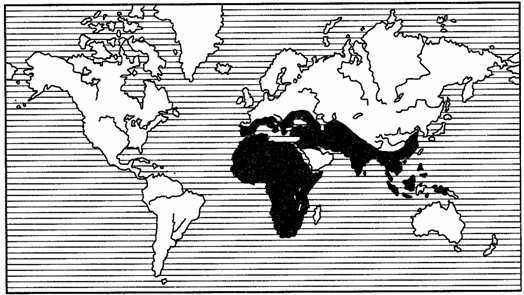
Рис. 120. Полирегиональный ареал жуков-навозников рода Onitis
(из В.Балтазари, 1963)
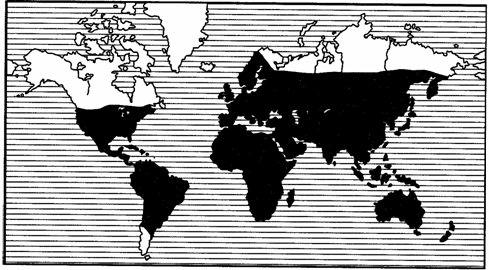
Рис. 121. Космополитный ареал жуков-навозников
рода Onthophagus (из В.Балтазари, 1963)
Между систематическим рангом группы и размером ареала существует корреляция: чем выше ранг группы, тем большую площадь она занимает. Следовательно, ареал рода больше ареала вида, ареал семейства больше ареала рода и т. д. Классы, как правило, распространены всесветно. Главными причинами, обусловливающими специфичность ареалов, являются экологическая валентность видов, их способность к расселению (вагильность), филогенетический возраст и скорость видообразования. Совокупность этих признаков и определяет величину ареала вида или их группы.
Формы ареалов, т. е. их очертания, столь же разнообразны, как и величина. Факторы, определяющие форму и величину ареала, чаще всего одни и те же. Очертания многих ареалов совпадают с ландшафтными или вообще с физико-географическими рубежами. В умеренных областях Северного полушария, а также в высоких широтах ареалы ряда видов вытянуты с запада на восток и значительно меньше — с севера на юг, т. е. носят зональный характер. Это объясняется особенностями распределения климатических условий, которые быстрее изменяются при продвижении с севера на юг, чем с запада на восток. В местах, далеко отстоящих друг от друга, но расположенных на одной широте, условия среды часто сходны.
Как показал Ю.И.Чернов (1975), наиболее точное совпадение ареала с конфигурацией зоны наблюдается в тех случаях, когда вид связан с зональным типом растительности. Зональные ареалы у «степняков» — джунгарского хомячка, кузнечика эверс-манна, степного таракана и др. Ареал лапландского подорожника Calcarius lapponicus совпадает с зоной тундры, мохноногого сыча — с таежной зоной; с широколиственными лесами связаны ареалы сонь — орешниковой, полчка и в некоторой степени лесной и садовой. Если вид и расселяется за пределы ландшафтной зоны, то он встречается лишь в экстразональных биотопах, не типичных для нового ландшафта.
В горных странах ареалы многих видов вытянуты вдоль хребтов и тоже укладываются в одну из вертикальных зон — лесную, субальпийскую и т. д. Объясняется это тем, что вдоль определенного склона хребта наблюдаются одинаковые условия, чаще климатические.
Вытягивание ареала, обусловленное экологическими факторами, может привести к образованию так называемых ленточных, или линейных, типов ареалов. Они свойственны многим околоводным или водным животным, особенно обитателям литорали. Аналогичную форму имеют ареалы пресноводных рыб. Хорошим примером такого ареала служит распространение среднеазиатских осетровых рыб из рода Pseudoscaphirhynchus, обитающих в Аму-дарье и Сырдарье.
Нередко встречаются и сопряженные ареалы. Их очертания определяются границами ареала другого вида. Это бывает в том случае, когда между видами существуют экологические взаимоотношения, которые обычно носят характер пищевой связи. Как правило, она устанавливается между паразитами и хозяевами и сожителями, хищниками и жертвами, животными-фитофагами и их кормовыми растениями: тигр связан с кабаном, куница — с белкой, соболь — с бурундуком (хищник — жертва), бабочка олеандровый бражник — с олеандром, жуки-нарывники Mylabris elegantissima — с песчаной акацией, африканский пальмовый орлан — с масличной пальмой (потребитель — кормовое растение). Монофаги встречаются только там, где есть кормовое растение. Однако ареалы последних больше, поскольку распространение животных зависит не только от наличия корма, но и от других факторов. Значительно реже наблюдается сопряжение ареалов паразита и хозяина. Паразиту часто требуется промежуточный хозяин, а то и два, и отсутствие одного из них, даже при наличии основного хозяина, препятствует распространению этого паразита.
Все рассмотренные выше ареалы, от узколокальных эндемичных до громадных, занимающих весь материк, относятся к сплошным ареалам.
Вместе с тем постоянно встречаются ареалы, разделенные на части, т. е. прерывистые, или дизъюнктивные. О них следует говорить в том случае, когда отдельные части ареала обособлены друг от друга в такой степени, что никакая связь между заселяющими их популяциями вида невозможна.
Ареал может прерываться по-разному. Бывает так, что в основном он сплошной, но встречаются обособленные маленькие участки его, которые принято называть островными местонахождениями. Ареалы подобного типа считаются расчлененными (фрагментированными). Они возникают и в силу естественных причин, но часто своим происхождением обязаны деятельности человека. Например, вырубка лесов приводит к исчезновению истинно лесных видов, сохраняющихся лишь в отдельных массивах.
Части дизъюнктивного ареала могут располагаться на одном материке, типы их разнообразны, но общим является то, что в этом случае между частями ареала нет преград в виде морских проливов или участков океана. На разделяющей территории природные условия тоже часто не отличаются от таковых в местах обитания видов, поэтому дать удовлетворительное объяснение причинам подобных разрывов довольно трудно. Типологические материковые дизъюнктивные ареалы могут различаться в зависимости от места расположения их частей. Выделяют, к примеру, евро-азиатские дизъюнктивные ареалы, когда одна их часть находится в Европе, другая — в Азии.
Как правило, части дизъюнктивного ареала связаны с одной ландшафтной зоной. Широко известными примерами такого рода являются амфиевро-азиатские неморальные ареалы амфибий — жерлянок рода Bombina, рыб — горчака и вьюна, птиц — голубой сороки и др. В широколиственных лесах Европы встречаются В. bombina и В. variegata, в Приморье их замещают В. orientalis, а в Китае — В. maxima (пример викариата на уровне видов). Горчак и голубая сорока в различных частях ареала представлены разными подвидами: в Европе живет горчак обыкновенный (Rhodeus sericeus sericeus), в бассейне Амура — горчак амурский (R. s. amurensis), в Испании — голубая сорока Cyanopica cyana cooki, а в Приморье — С. с. cyanus (рис. 122).
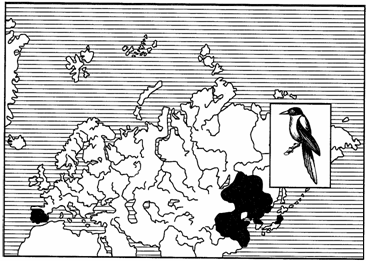
Рис. 122. Амфиевро-азиатский неморальный ареал голубой сороки
Различают также арктоальпийский и бореомонтанный типы дизъюнктивного ареала. В первом случае основная часть ареала (зональная) находится в тундре, другая — в альпийском поясе гор Европы (рис. 123).
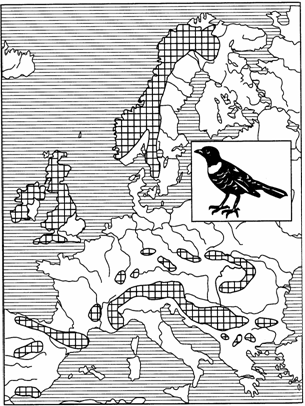
Рис. 123. Арктоальпийский ареал белозобого дрозда (из Г.Делаттина, 1967)
Особый и нередко встречающийся тип дизъюнкций представляют горные разрывы ареалов, когда отдельные их части находятся в верхних поясах горных хребтов и отделены друг от друга более или менее широкими долинами и межгорными понижениями. К примеру, серна — высокогорный копытный зверь — в каждом горном массиве имеет изолированный ареал. Популяции ее в Пиренеях, Альпах, на Балканах, горах Тавр в Малой Азии и на Кавказе не связаны друг с другом (рис. 124).
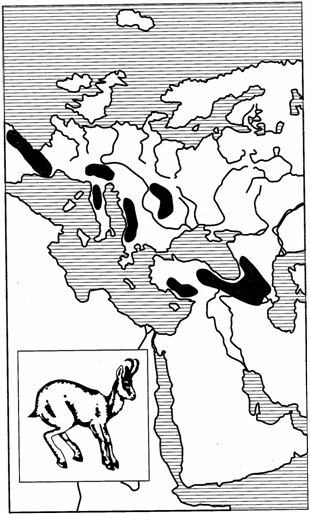
Рис. 124. Горный разрыв ареала серны (из И.К.Лопатина, 1980) 336
Аналогичный горный разрыв характерен для ареалов видов одного рода, например горных козлов Сарга. Последние заселяют горы Южной Европы, Малой и Средней Азии, Гималаев, Южной Сибири. Подобные ареалы характерны для высокогорных насекомых, например для представителей уже упоминавшегося рода жуков-листоедов Oreomela, объединяющего более 50 видов, причем только на Тянь-Шане их 28, в Джунгарии и Синьцзяне — 11, в Тибете и Цинхае (КНР) — 9. Каждому хребту свойственны свои виды, а межгорные долины и ущелья являются непреодолимыми преградами для их расселения, поскольку жуки не опускаются ниже пояса криволесья и чаще населяют только альпийский пояс хребтов.
Причины материковых разрывов ареалов разнообразны. Чаще разъединение объясняется фрагментацией сплошного ареала, вызванной изменением климата, конкуренцией более приспособленных форм, геоморфологическими перестройками или, наконец, деятельностью человека (вырубка лесов, мелиорация болот, распашка целины и т.п.). В любом из этих случаев можно допустить либо вымирание отдельных популяций, либо вытеснение их другими, более приспособленными к новым условиям видами, что иногда подтверждается палеонтологическими находками в области разрыва.
В Северном полушарии ряд дизъюнкций ареалов обусловлен событиями плейстоцена, в связи с чем они получили название ледниковых. Допускается, что некогда сплошные зональные ареалы Северного полушария были разорваны наступившими льдами. Там же, где покровного обледенения не было, климат настолько ухудшился, что некоторые популяции вымерли, а сохранившиеся в наиболее благоприятных районах не смогли расселиться и восстановить свой прежний ареал. Эти климатические изменения не коснулись теплолюбивых неморальных видов, уцелевших в убежищах, находившихся в юго-западной Европе и на юге Дальнего Востока (они там живут и в современную эпоху). Климат же Сибири изменился настолько, что расселение через нее стало невозможным. События ледникового периода вызвали также разрывы ареалов арктоальпийского и бореомонтанного типов. Во время четвертичного покровного оледенения климат Северной Европы был холодным, и характерная для него арктическая (тундровая), а также бореальная (таежная) фауна широко распространилась к югу. Достаточно сказать, что, по данным палеонтологии, заяц-беляк, песец и рысь жили в то время даже в Крыму. В альпийской зоне южных гор, в свою очередь, развивалось местное оледенение и ледниковые языки опускались далеко в долины. После таяния и отступания материковых льдов холодолюбивые виды, обитавшие на окраинах ледника, мигрировали на север и заняли здесь освободившиеся ото льда пространства. Популяции некоторых видов при сокращении горного оледенения отступали вместе с ледниками в самые верхние пояса хребтов, где климат оставался холодным. Между горами Центральной и Южной Европы, с одной стороны, и тайгой или тундрой — с другой, образовались разрывы ареалов. Северная часть нынешнего ареала рассматривается как новый (по времени) ареал, возникший в результате активного расселения животных, а южная (горная) — как реликтовая. Таким образом, заяц-беляк и тундряная куропатка в Альпах являются ледниковыми реликтами.
Горные разрывы ареалов возникли в результате сложных геоморфологических процессов, происходивших в определенных частях земного шара.
Особый тип дизъюнктивных ареалов составляют ареалы, у которых между их частями находятся морские пространства. Подобные океанические дизъюнкции ареалов характерны для надвидо-вых таксонов — родов, семейств, отрядов и т. д. В пределах Северного полушария к данному типу ареалов относятся циркумполизональ-ные, циркумполярные, циркумбореальные и циркумтемператные (расположены по всей умеренной зоне), а также амфиатланти-ческие ареалы (рис. 125, 126). Части последних располагаются на материках Евразии и Северной Америки в соответствующих зонах, а циркумполизональные и панголарктические охватывают все зоны материков.
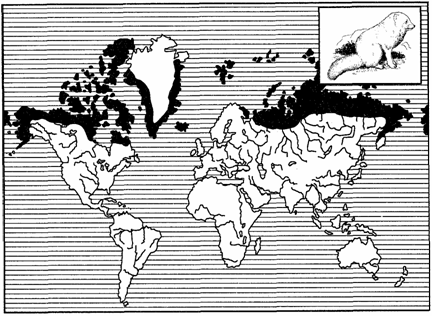
Рис. 125. Циркумполярный ареал песца (из В. Г. Гептнера, 1936)
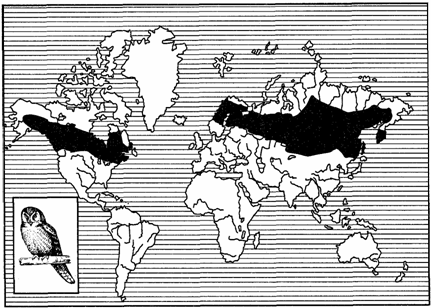
Рис. 126. Циркумбореальный ареал ястребиной совы
(из В. Г. Гептнера, 1936)
Очень обычны формы европейско-североамериканского и ази-атско-американского разрывов. В этих случаях между ареалами родственных наземных и пресноводных таксонов находятся водные преграды — Атлантический океан, Берингов пролив или даже северная часть Тихого океана.
Примерами океанических разрывов ареалов может служить распространение тапиров (рис. 127) в тропических частях Южной Америки и юго-восточной Азии, сумчатых млекопитающих (рис. 128), жаб-свистунов (рис. 129) в Австралии и Южной Америке и др.
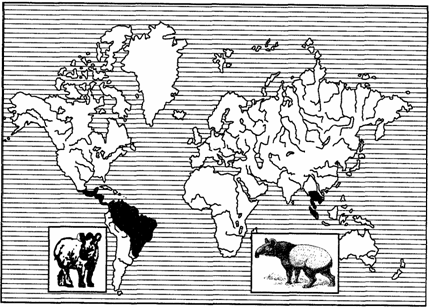
Рис. 127. Дизъюнктивный ареал тапиров (из И.К.Лопатина, 1980)
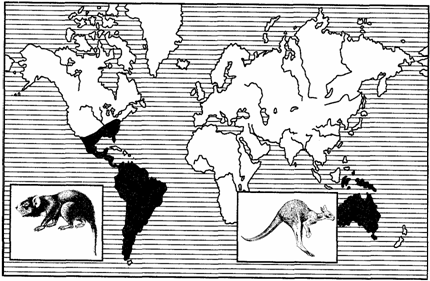
Рис. 128. Дизъюнктивный ареал сумчатых млекопитающих
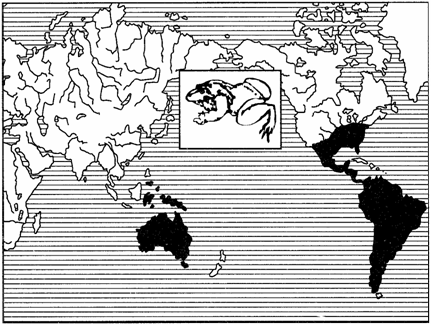
Рис. 129. Дизъюнктивный ареал жаб-свистунов (из И.К.Лопатина, 1980)
Среди океанических разрывов замечательны случаи биополярного и амфибореального распространения (рис. 130): соответствующие группы животных встречаются в приполярных или умеренно холодных частях Земли и отсутствуют в тропиках и субтропиках.
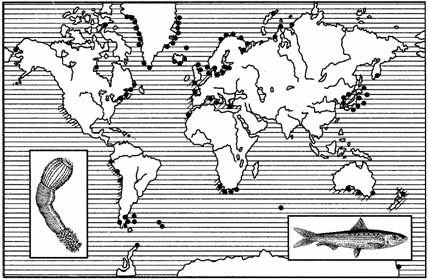
Рис. 130. Биополярное распространение червей-приапулид
(обозначено точками) и амфибореальное распространение
сардин (обозначено штрихом)
Причины многих океанических разрывов ареалов не установлены, особенно когда нет палеонтологических находок на территориях, где те или иные группы животных не встречаются. Предложено несколько теорий, объясняющих происхождение подобных ареалов (теории мостов суши, оттесненных реликтов и др.).
Ареалы близких видов располагаются рядом, а иногда могут накладываться друг на друга (перекрывание ареалов). Так, ареалы зайца-беляка и зайца-русака в Северной Европе на большом протяжении расположены на одной и той же территории. Бывает, что ареал одного вида, как правило, небольшой величины, охвачен гораздо большим ареалом другого вида. Из-за различных требований, предъявляемых видами к окружающей среде, перекрывание их ареалов — явление нередкое. Оно получило название симпатрии.
Иная картина наблюдается при изучении ареалов подвидов. В данном случае ареалы очень часто прилегают друг к другу, но никогда не перекрываются. Каждый подвид населяет определенную область видового ареала (подвиды аллопатричны), и подвиде -вые ареалы замещают друг друга, или викарируют. Примером географического викариата служит распространение подвидов колхидского фазана на территории от Кавказа до Тихого океана. Викарируют не только подвиды, но и близкие виды. Общеизвестно, что европейскому зубру (Bison bonasus) соответствует в Северной Америке викарный вид бизон (Bison bison). В рассмотренных случаях замещающие формы не только близки по происхождению, но и занимают одинаковые биотопы. Такой тип викариата назван систематическим. Если же одинаковые биотопы в разных странах населены различными по систематическому положению и происхождению видами, конвергентно сходными между собой из-за одинакового образа жизни, то пользуются термином «экологический викариат». Примером экологического викариата является замещение европейского крота в Африке златокротом (другое семейство), а в Австралии — сумчатым кротом (другой подкласс).
Величина ареалов и их границы находятся в тесной взаимосвязи. Если размеры ареалов очень часто зависят от экологической валентности населяющих их видов, то значение экологических факторов, определяющих границы ареалов, еще более велико. Эти границы нередко обусловлены определенной комбинацией факторов среды или даже их одним градиентом. Особое значение имеют температура, освещение, питание, характер субстрата. Для наземных животных существенную роль, кроме того, играет влажность воздуха, для водных — химизм воды, содержание кислорода и т. д.
Существует разделение границ на климатические, ландшафтные, биоценотические. Их можно назвать экологическими границами, поскольку они определяются целой совокупностью факторов. С другой стороны, известно немало случаев, когда граница остается стабильной, и расселения не происходит, несмотря на то что условия среды внутри ареала и за его пределами однородны и вполне благоприятны для вида. Подобные явления объясняются историческими причинами и указывают на то, что в прошлом условия среды по обе стороны границ были различными. Границы ареалов, происхождение которых обусловлено минувшими событиями и не оправдывается экологической обстановкой настоящего времени, называются историческими или реликтовыми.
Биоценотические границы определяются взаимоотношениями разных видов в сообществе. Эти взаимоотношения носят разный характер (симбиоз, комменсализм, конкуренция и т. п.). Биологическое соперничество — одна из причин стабильности границ ареалов двух конкурирующих видов. Так, например, обстоит дело с ареалами двух близких видов — лесной куницы и соболя на Урале и в Западной Сибири.
Однако стабильность границ — явление относительное и временное, тем более в настоящий период существования животного мира, когда в процесс расширения и сокращения ареалов вмешивается человек.

Обсуждение Основы зоологии и зоогеографии
Комментарии, рецензии и отзывы