Происхождение многоклеточных
Происхождение многоклеточных
В основе большинства представлений о возникновении многоклеточных лежит давнее убеждение, что они произошли от колоний Protozoa и что, следовательно, тело одноядерного простейшего в морфологическом отношении отвечает отдельной клетке многоклеточного животного. При этом полагают, что в процессе эволюции постепенно развилась новая индивидуальность многоклеточного организма, резкое подчинение ей и подавление индивидуальности отдельных клеток. Иными словами, метазоон по сравнению с простейшим признается индивидом высшего порядка. Колониальные гипотезы, таким образом, в полном соответствии с клеточной теорией рассматривают клетку как элементарную структурную единицу, позволяющую сравнивать и анализировать организацию всех Protozoa, Metazoa и Metaphyta (многоклеточных растений).
Гипотеза гастреи Э. Геккеля. Первую гипотезу о колониальном происхождении многоклеточных — гипотезу «гастреи» — предложил Э. Геккель. В основу этой гипотезы, которую он разрабатывал с начала 70-х гг. XIX в., легла идея о гомологии зародышевых листков у всех многоклеточных животных, высказанная впервые Т. Гексли. Ко времени появления гипотезы гастреи учение о зародышевых листках сделало большие успехи благодаря работам Т. Гексли, К. Ф. Вольфа, К. Бэра и др. Геккель опирался на достижения эмбриологии своего времени и в частности на исследования А. О. Ковалевского.
Важнейшим «орудием» при создании теории гастреи был биогенетический закон, обоснованный почти одновременно Ф. Мюллером и Геккелем в 60-е гг. XIX в. Согласно Геккелю, «онтогения представляет собой краткое повторение филогении, механически обусловленное функциями наследственности и приспособляемости» (Haeckel, 1874). Он рассматривал так называемые первичные зародышевые листки — эктодерму и энтодерму гаструлы как проявление в онтогенезе соответственных примитивных органов первобытных предков. Всем прочим начальным стадиям онтогенеза Геккель также приписывал абсолютное рекапитуля-ционное значение. Все характерные стадии дробления '(рис. 12) отвечают, по мнению Геккеля, сходным стадиям филогенеза. Так, яйцу, или цистуле, соответствует одноклеточный предок Cytaea, стадии морулы — предковая форма «морея» и т. п. Особенно важной и широко распространенной в животном мире рекапитуляцией (т. е. повторением филогенеза в онтогенезе) Геккель считал двухслойную зародышевую стадию — гаструлу. Общего гипотетического прародителя всех Metazoa он создал по ее образу и подобию.
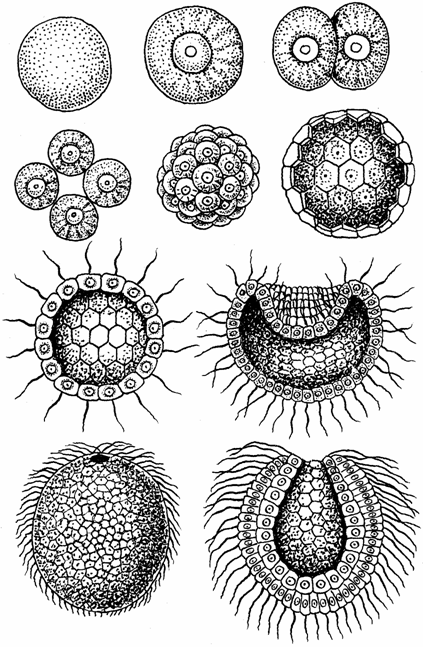
Рис.12. Стадии эмбрионального развития кораллового
полипа (по Геккелю)
Первой филогенетической стадией, по Геккелю, был одноклеточный амебообразный организм. От него произошли все ани-мально питающиеся организмы. Колония одинаковых амебоидных клеток дала затем начало «морее» — плотному шарообразному организму, рекапитуляцию которого в онтогенезе представляет морула. Путем накопления в центре морей жидкости или студенистого вещества, вытеснившего клетки на периферию, постепенно сформировалась свободноплавающая «бластея» (в онтогенезе ей отвечает бластула). Клетки бластеи сперва были покрыты псевдоподиями, которые позднее приобрели способность быстро двигаться и изгибаться и превратились в жгуты, служащие для плавания.
Следующей очень важной стадией была гастрея, образовавшаяся из бластеи путем выпячивания (инвагинации) стенки тела на переднем полюсе. Наружный клеточный слой гастреи был снабжен жгутиками и сохранил функцию движения, внутренний слой стал кишечным. В центральной, кишечной, полости, сообщавшейся ртом с наружной средой, происходило переваривание заглоченной добычи. Два эпителиальных слоя гастреи — эктодерма и энтодерма — представляли собой первичные органы, из которых у потомков гастреи возникли все их органы и ткани.
Современных кишечнополостных и губок Геккель рассматривал как мало изменившихся потомков гастреи, а стадию гаструлы — как рекапитуляцию гастреи.
Все многоклеточные, согласно Геккелю, в отличие от простейших имеют монофилетическое происхождение и развились от одной прародительской формы — гастреи, произошедшей, в свою очередь, от одноклеточных предков. От гастреи все Metazoa унаследовали первичный кишечник и два первичных зародышевых листка, их ткани представляют собой дериваты этих двух листков.
Свою гипотезу Геккель характеризовал как попытку улучшить филогенетические основы естественной классификации и выяснить пути развития главных систематических групп животного мира. Действительно, из гипотезы гастреи вытекал ряд существенных выводов для систематики, сравнительной анатомии, эмбриологии и гистологии. Однако самым важным следствием появления гипотезы гастреи было разрушение учения о типах Кювье, все еще господствовавшего в зоологии того времени.
Из гипотезы Геккеля следует, что истинные гомологии органов и их систем возможны у всех потомков гастреи, т. е. у представителей разных типов, тогда как теорией типов эта возможность отрицалась. Так как гаструла у всех Metazoa гомологична, то гомо-логичен всегда и кишечник. Гомологичны, далее, у всех Metazoa и кожные покровы, поскольку всегда имеется слой эпидермиса, служащий источником для развития других кожных слоев — кутикулы, железистых образований и пр. — и отвечающий эктодерме гастреи. Нервная система всегда развивается из эктодермы и гомологична во всех группах животных. Геккель видел также основания для гомологизации выделительных органов, целома и кровеносной системы у тех животных, у которых они имеются. Для органов чувств, скелета и сердца он считал общую гомологию неприемлемой и полагал, что все эти органы развились в разных группах независимо. Он признавал достоверным различное происхождение ротового отверстия в разных группах многоклеточных. Бластопор гаструлы, гомологичный рту гастреи, сохраняется во взрослом состоянии у кишечнополостных, у губок (в виде устья) и у низших червей. Рот иглокожих, членистоногих и позвоночных, по Геккелю, есть новообразование.
Таким образом, Геккель признавал широкие возможности конвергентного развития различных важных особенностей в строении животных.
Он считал первичной тканью однослойный жгутиковый эпителий, а все прочие ткани — вторичными производными эпителия. Эктодерму и энтодерму Геккель считал первичными зародышевыми листками. Мезодерма же, по его мнению, возникла в процессе эволюции гораздо позднее, так как в онтогенезе она всегда образуется из эктодермы и энтодермы и, по существу, даже не представляет единого листка, а имеет двойственную природу, слагаясь из пластинок, развившихся независимо из кожно-мус-кульной и кишечномускульной пластинок.
Так как мезодерма всегда развивается из парных зачатков, то, по Геккелю, у разных групп животных она имеет общее происхождение и может считаться гомологичной. Первичные зародышевые листки у низших — губок и кишечнополостных — в отличие от таковых у высших типов сохраняются как первичные органы, подобно тому, как это имело место у гипотетической гастреи.
Гипотеза Геккеля долгое время была господствующей, некоторые крупные зоологи придерживаются ее и теперь. Ее положительная роль в зоологии была чрезвычайно велика, так как она показала единство и общность происхождения всех многоклеточных и тем способствовала прогрессу дарвинизма.
Однако гипотеза гастреи страдает существенными недостатками, которые не укрылись уже от некоторых современников Геккеля и давали повод для ее резкой критики.
Действительно, гипотеза гастреи не согласуется со многими данными зоологии и должна уступить место более совершенной концепции. Впрочем, учение о протозойных колониальных предках Metazoa, лежащее в основе обобщений Геккеля, целиком сохраняет свое значение и поныне. Вторым «рациональным» зерном гипотезы гастреи следует считать учение о бластее, которое без особенных изменений было воспринято авторами некоторых других колониальных гипотез.
Известный русский эмбриолог В. В. Заленский (1874) подробно рассмотрел первые стадии эмбрионального развития различных животных с точки зрения соответствия их теории гастреи. Главнейшим моментом в онтогенезе животных он считал первую дифференциацию зародышевых листков. Общий ход рассуждении В. В. Заленского был таков. В типичных случаях у низших многоклеточных после дробления и стадии морулы формируется двуслойная бескишечная форма — планула. Если же образуется полая шаровидная бластула, то затем в ее полости появляются энтодермальные клетки и возникает стадия (дибластула), вполне сравнимая с планулой, так как она, по существу, тоже имеет два зародышевых листка и лишена эпителиальной кишки. В тех случаях, когда путем впячивания образуется гаструла с мешковидной кишкой и ртом, мы, на взгляд Заленского, имеем вторично измененное развитие, обеспечивающее очень раннее появление кишечника и характеризующееся выпадением стадии планулы. Поэтому Заленский думал, что общий предок Metazoa, скорее, должен был обладать признаками планулы, нежели гастреи. Заленский, фактически, был предшественником И. И. Мечникова, выдвинувшего известную гипотезу фагоцителлы.
Гипотеза фагоцителлы И.И.Мечникова. Подобно Заленскому, И. И. Мечников подверг гипотезу гастреи резкой критике. В частности, он заметил, что идентичности первичной гаструлы у всех Metazoa, принимаемой Геккелем, в действительности не существует. У разных животных эта стадия имеет различные особенности и развивается по-разному, что далеко не всегда можно объяснить вторичными причинами. Истинные двуслойные, инвагинационные гаструлы, как этого требует теория гастреи, в действительности крайне редки. В законченном виде гипотеза фагоцителлы И. И. Мечникова изложена в заключительной главе его монографии «Эмбриологические исследования над медузами» (1886).
Будучи сторонником колониального происхождения, И. И. Мечников, как и Геккель, видел отдаленных предков многоклеточных в жгутиконосцах с животным питанием.
Мечников считал инвагинацию, путем которой образуется гаструла, вторичным способом образования энтодермы, возникшим в результате длительной и сложной эволюции.
Гипотеза И. И. Мечникова состоит в следующем. Первичный метазоон был шаровидным и имел сначала однослойное строение. Иными словами, признается бластея, и в этом — совпадение с гипотезой Геккеля.
Поскольку у Metazoa полость дробления обычно появляется очень рано и зародыш быстро превращается в бластулу, предком многоклеточных Мечников считал бластулообразную колонию жгутиконосцев. Он полагал, что тотальное дробление многоклеточных следует выводить из деления жгутиконосцев: первые меридиональные деления дробящегося яйца представляют собой наследие от флагелятных предков, поскольку для жгутиконосцев характерно именно продольное деление. Исходную шарообразную форму колонии Мечников пытается также объяснить исходя из продольного деления жгутиконосцев. Если деление клетки все время происходит продольно, то получается пластинка, но если третье деление изменится, станет поперечным, то в результате возникает шаровидная колония клеток. Именно такое изменение направления деления и произошло в филогенезе. Таким образом, предком Metazoa была колония, у которой направления деления чередовались в трех координатных плоскостях. Мечников думал, что образование двуслойной стадии произошло не путем впячива-ния, а путем иммиграции — внедрения отдельных клеток в полость бластулы, в результате чего образовался зачаток энтодермы. Эволюционные истоки такой иммиграции он видел в явлении фагоцитоза.
Питание первичного метазоона, по Мечникову, совершалось теми же клетками, которые служили для движения, т. е. жгутиковыми клетками путем внутриклеточного пищеварения (фагоцитоза). И. И. Мечников предположил, что перегруженные пищей клетки легко теряли жгутик и уходили в полость организма, затем они снова могли выходить на поверхность и формировать жгутик. Так получилась первая факультативная дифференциация на наружный слой клеток — «кинобласт» — и на внутреннюю клеточную массу — «фагоцитобласт». Эта дифференциация затем закрепилась в эволюции, и образовался компактный организм — паренхимелла, моделью которого он считал личинку губок — паренхимуллу. Позднее Мечников назвал этот организм фагоцител-лой. Это и был общий предок многоклеточных животных.
Дальнейшая судьба фагоцителлы такова. Некоторые ее потомки перешли к сидячему образу жизни и дали начало губкам. Другие стали ползать, приобрели билатеральную симметрию и ротовое отверстие. От них произошли бескишечные плоские черви турбеллярии, у которых еще нет кишечника и пищеварение совершается в лакунах паренхимы и в блуждающих клетках — фагоцитах. Третьи, сохранив плавающий образ жизни, приобрели рот, испытали эпителизацию фагоцитобласта и превратились в первичных кишечнополостных — родоначальников сидячих полипов.
Таким образом, гипотеза И. И. Мечникова объясняла с эволюционной точки зрения все главные этапы онтогенеза Metazoa и предлагала новые филогенетически обоснованные представления о первичных зародышевых листках и их дальнейшей эволюции. На этой основе Мечников нарисовал вполне правдоподобную гипотетическую картину эволюционного становления Metazoa и первых этапов их филогенетического развития, картину, которая хорошо объясняет множество эмбриологических и сравнительно-анатомических факторов, непонятных с точки зрения других гипотез.
А. А. Захваткин в 1949 г. выдвинул гипотезу о происхождении многоклеточных от колониальных жгутиконосцев на основе па-линтомии — особой формы бесполого размножения путем последовательных делений клетки без стадий роста получающихся дочерних клеток. Такое деление является, по его мнению, прообразом дробления яйца у Metazoa.
Другой путь эволюционного формирования Metazoa предложил А. В. Иванов в конце 60-х гг., считавший, что гипотетические исходные колонии жгутиконосцев не были палинтомическими и вообще мало отличались от шаровидных колоний современных во-ротничковых жгутиконосцев.
Иванов принимает за основу теорию фагоцителлы Мечникова. Однако прообразом фагоцителлы он считает не личинку губок, а примитивное плоское многоклеточное трихоплакс, являющееся единственным представителем типа Пластинчатые (Placozoa). Схема возникновения многоклеточных, согласно Иванову, представлена на рис. 13.
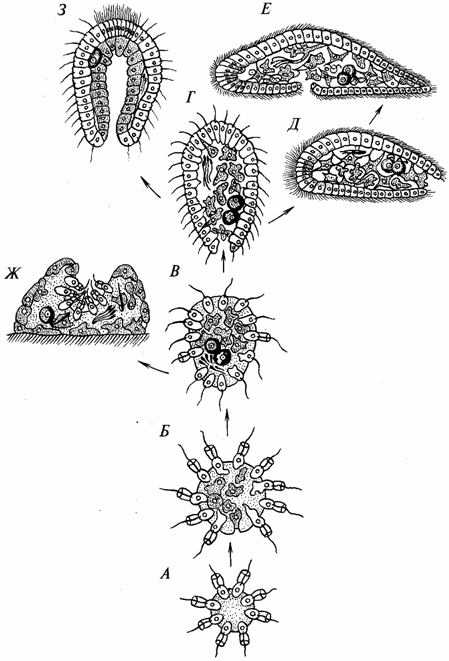
Рис. 13. Главнейшие предполагаемые стадии филогенеза Metazoa
по А. В. Иванову:
1 — колония жгутиконосцев; Б — миграция клеток жгутиконосцев внутрь; В — ранняя фагоцителла; Г — поздняя фагоцителла; Д — первичная турбеллярия — появление рта и билатеральной симметрии; Е — примитивная бескишечная турбеллярия — усиление дифференциации клеток, смещение рта на брюшную сторону; Ж — примитивная губка — переход к сидячему образу жизни, замена локомоторной функции киноцитов гидрокинетической; 3 — первичное кишеч-нополостное типа гастреи — образование рта, эпителизация фагоцитобласта
Поскольку в эмбриогенезе низших многоклеточных двуслойных зародыш образуется чаще путем иммиграции, большинство зоологов считают, что именно таким путем и шло преобразование шаровидной колонии жгутиконосцев в первый многоклеточный организм. При этом у предковых форм многоклеточных образование двух клеточных слоев сопровождалось специализацией клеток и колония жгутиконосцев превратилась в единый многоклеточный организм. Наружный слой сохранил двигательную и чувствительную функции, а внутренний — пищеварительную и половую.

Обсуждение Основы зоологии и зоогеографии
Комментарии, рецензии и отзывы