Страница 41
Посттрансляционные преобразования белков. Синтезированные в ходе трансляции пептидные цепи на основе своей первичной структуры приобретают вторичную и третичную, а многие—и четвертичную организацию, образуемую несколькими пептидными цепями. В зависимости от функций, выполняемых белками, их аминокислотные последовательности могут претерпевать различные преобразования, формируя функционально активные молекулы белка.
Многие мембранные белки синтезируются в виде пре-белков, имеющих на N-конце лидерную последовательность, которая обеспечивает him узнавание мембраны. Эта последовательность отщепляется при созревании и встраивании белка в мембрану. Секреторные белки также имеют на N-конце лидерную последовательность, которая обеспечивает их транспорт через мембрану.
Некоторые белки сразу после трансляции несут дополнительные аминокислотные про-последовательности, определяющие стабильность предшественников активных белков. При созревании белка они удаляются, обеспечивая переход неактивного пробелка в активный белок. Например, инсулин вначале синтезируется как пре-проинсулин. Во время секреции пре-последовательность отщепляется, а затем проинсулин подвергается модификации, при которой из него удаляется часть цепи и он превращается в зрелый инсулин.
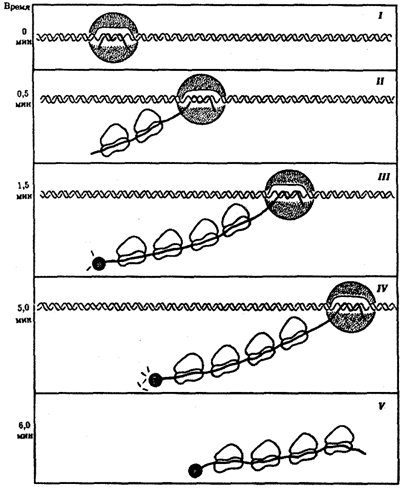
Рис. 3.41. Транскрипция, трансляция и деградация мРНК у прокариот:
I — РНК-полимераза связывается с ДНК и начинает синтезировать мРНК в направлении 5' → 3';
II — по мере продвижения РНК-полимеразы к 5'-концу мРНК прикрепляются рибосомы, начинающие синтез белка;
III — группа рибосом следует за РНК-полимеразой, на 5'-конце мРНК начинается ее деградация;
IV —процесс деградации протекает медленнее, чем транскрипция и трансляция;
V — после окончания транскрипции мРНК освобождается от ДНК, на ней продолжается трансляция и деградация на 5'-конце
Формируя третичную и четвертичную организацию в ходе посттрансляционных преобразований, белки приобретают способность активно функционировать, включаясь в определенные клеточные структуры и осуществляя ферментативные и другие функции.
Рассмотренные особенности реализации генетической информации в прои эукариотических клетках обнаруживают принципиальное сходство этих процессов. Следовательно, механизм экспрессии генов, связанный с транскрипцией и последующей трансляцией информации, которая зашифрована с помощью биологического кода, сложился в целом еще до того, как были сформированы эти два типа клеточной организации. Дивергентная эволюция геномов прои эукариот привела к возникновению различий в организации их наследственного материала, что не могло не отразиться и на механизмах его экспресии.
Постоянное совершенствование наших знаний об организации и функционировании материала наследственности и изменчивости обусловливает эволюцию представлений о гене как функциональной единице этого материала.
3.4.3.3. Ген — функциональная единица наследственного материала. Взаимосвязь между геном и признаком
Долгое время ген рассматривали как минимальную часть наследственного материала (генома), обеспечивающую развитие определенного признака у организмов данного вида. Однако каким образом функционирует ген, оставалось неясным. В 1945 г. Дж. Бидлом и Э. Татумом была сформулирована гипотеза, которую можно выразить формулой «Один ген — один фермент». Согласно этой гипотезе, каждая стадия метаболического процесса, приводящая к образованию в организме (клетке) какого-то продукта, катализируется белком-ферментом, за синтез которого отвечает один ген.
Позднее было показано, что многие белки имеют четвертичную структуру, в образовании которой принимают участие разные пептидные цепи. Например, гемоглобин взрослого человека включает четыре глобиновых цепи — 2α и 2β, кодируемые разными генами. Поэтому формула, отражающая связь между геном и признаком, была несколько преобразована: «Один ген — один полипептид».
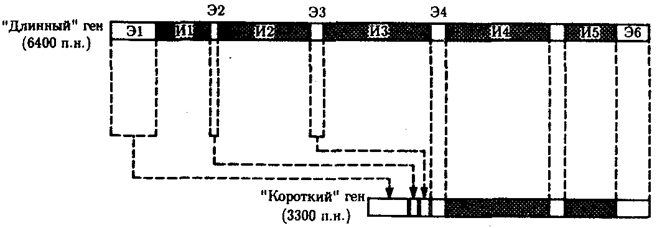
Рис. 3.42. Две формы гена box (cob) митохондрий дрожжей,
кодирующего цитохром b
«Короткий» ген box образуется в результате удаления из «длинного» гена первых трех интронных участков и сплайсинга первых четырех экзонов; И — интрон, Э — экзон; п.н. — пары нуклеотидов
Изучение химической организации наследственного материала и процесса реализации генетической информации привело к формированию представления о гене как о фрагменте молекулы ДНК, транскрибирующемся в виде молекулы РНК, которая кодирует аминокислотную последовательность пептида или имеет самостоятельное значение (тРНК и рРНК).
Открытия экзон-интронной организации эукариотических генов и возможности альтернативного сплайсинга показали, что одна и та же нуклеотидная последовательность первичного транскрипта может обеспечить синтез нескольких полипептидных цепей с разными функциями или их модифицированных аналогов. Например, в митохондриях дрожжей имеется ген box (или cob), кодирующий дыхательный фермент цитохром b. Он может существовать в двух формах (рис. 3.42). «Длинный» ген, состоящий из 6400 п. н., имеет 6 экзонов общей протяженностью 1155 п.н. и 5 интронов. Короткая форма гена состоит из 3300 п.н. и имеет 2 интрона. Она фактически представляет собой лишенный первых трех интронов «длинный» ген. Обе формы гена одинаково хорошо экспрессируются.
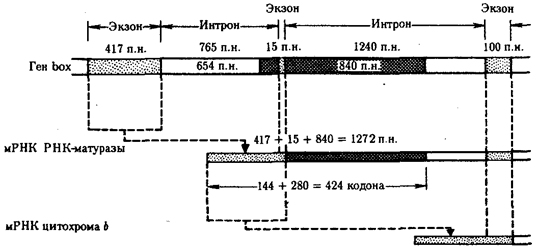
После удаления первого интрона «длинного» гена box на основе объединенной нуклеотидной последовательности двух первых экзонов и части нуклеотидов второго интрона образуется матрица для самостоятельного белка — РНК-матуразы (рис. 3.43). Функцией РНК-матуразы является обеспечение следующего этапа сплайсинга — удаление второго интрона из первичного транскрипта и в конечном счете образование матрицы для цитохрома b.

Обсуждение Биология Том 1
Комментарии, рецензии и отзывы