Страница 7
При физической (механической) изоляции препятствием к скрещиванию являются различия в структуре органов размножения или просто разница в размерах тела. У растений такая форма изоляции возникает при приспособлении цветка к определенному виду опылителей.
Описанные формы изоляции, особенно в начальный период их действия, снижают, но не исключают полностью межпопуляционные скрещивания.
Генетическая (репродуктивная) изоляция создает более жесткие, иногда непреодолимые барьеры скрещиваниям. Она заключается в несовместимости гамет, гибели зигот непосредственно после оплодотворения, стерильности или малой жизнеспособности гибридов.
Иногда разделение популяции сразу начинается с генетической изоляции. К этому приводят полиплоидия или массивные хромосомные перестройки, резко изменяющие хромосомные наборы гамет мутантов по сравнению с исходными формами. Полиплоидия распространена среди растений (рис. 11.2). Разные виды плодовой мухи нередко различаются хромосомными перестройками. Гибриды от скрещивания близкородственных форм со сниженной жизнеспособностью известны для серой и черной ворон. Указанный фактор изолирует популяции этих птиц в Евразии (рис. 11.3). Чаще генетическая изоляция развивается вторично вследствие углубления морфологических различий организмов из популяций, длительно разобщенных другими формами изоляции — географической, биологической. В первом случае генетическая изоляция предшествует дивергенции признаков и начинает процесс видообразования, во втором — она его завершает.


Рис.11.2. Растения и наборы хромосом в соматических клетках Solanum nigrum:
а—г: 36, 72, 108 и 144 хромосомы
Изоляция в процессе видообразования взаимодействует с другими элементарными эволюционными факторами. Она усиливает генотипические различия, создаваемые мутационным процессом и генетической комбинаторикой. Возникающие благодаря изоляции внутривидовые группировки отличаются по генетическому составу и испытывают неодинаковое давление отбора.
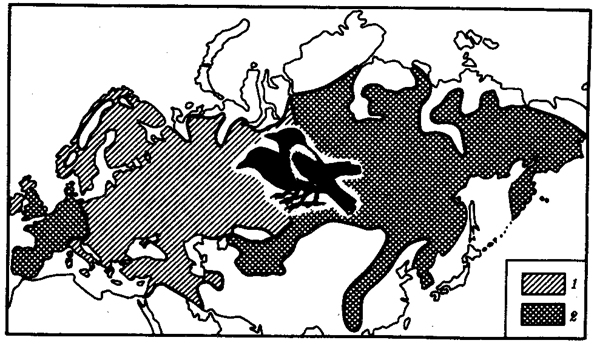
Рис. 11.3. Сниженная жизнеспособность гибридов как фактор разделения популяций серой и черной ворон:
1 — ареал серой вороны, 2 — ареал черной вороны
11.4. ЕСТЕСТВЕННЫЙ ОТБОР
В природных популяциях организмов, размножающихся половым способом, существует большое разнообразие генотипов и, следовательно, фенотипов. Благодаря индивидуальной изменчивости в условиях конкретной среды обитания приспособленность разных генотипов (фенотипов) различна. В эволюционном контексте приспособленность определяют как произведение жизнеспособности в данной среде, обусловливающей большую или меньшую вероятность достижения репродуктивного возраста, на репродуктивную способность особи. Различия между организмами по приспособленности, оцениваемой передачей аллелей следующему поколению, выявляются в природе с помощью естественного отбора. Главный результат отбора заключается не просто в выживании более жизнеспособных, а в относительном вкладе таких особей в генофонд дочерней популяции.
Необходимой предпосылкой отбора является борьба за существование — конкуренция за пищу, жизненное пространство, партнера для спаривания. Естественный отбор происходит на всех стадиях онтогенеза организмов. На дорепродуктивных стадиях индивидуального развития, например в эмбриогенезе, преобладающим механизмом отбора служит дифференциальная (избирательная) смертность. В конечном итоге отбор обеспечивает дифференциальное (избирательное) воспроизведение (размножение) генотипов. Благодаря естественному отбору аллели (признаки), повышающие выживаемость и репродуктивную способность, накапливаются в ряду поколений, изменяя генетический состав популяций в биологически целесообразном направлении. В природных условиях естественный отбор осуществляется исключительно по фенотипу. Отбор генотипов происходит вторично через отбор фенотипов, которые отражают генетическую конституцию организмов.
Как элементарный эволюционный фактор естественный отбор действует в популяциях. Популяция является полем действия, отдельные особи — объектами действия, а конкретные признаки — точками приложения отбора.
Эффективность отбора по качественному и количественному изменению генофонда популяции зависит от величины давления и направления его действия. Величину давления отбора выражают коэффициентом отбора S, который характеризует интенсивность устранения из репродуктивного процесса или сохранения в нем соответственно менее или более приспособленных форм по сравнению с формой, принятой за стандарт приспособленности. Так, если некий локус представлен аллелями A1 и А2, то популяция по генотипам делится на три группы: A1A1; A1A2; A2A2. Обозначим приспособленность этих генотипов W0, W1, W2. Выберем в качестве стандарта первый генотип, относительная приспособленность которого максимальна и равна 1. Тогда приспособленности других генотипов составят доли этого стандарта:
AlA1 A1A2 A2A2
W0 W1 W2
или W0/W0 = 1, W1/W0=l—S1, W2/W0=l—S2.
Величины S1 и S2 означают пропорциональное снижение в очередном поколении воспроизводства генотипов A1A2 и А2А2 сравнительно с генотипом AlA1.
Отбор особенно эффективен в отношении доминантных аллелей при условии их полного фенотипического проявления и менее эффективен в отношении рецессивных аллелей, а также в условиях неполной пенетрантности. На результат отбора влияет исходная концентрация аллеля в генофонде. При низких и высоких концентрациях отбор происходит медленно. Изменение доли доминантного аллеля в сравнении с рецессивным при коэффициенте отбора 0,01 приведено ниже.
Возрастание, %

Обсуждение Биология Том 2
Комментарии, рецензии и отзывы