Страница 131
Репрессор обладает свойством специфически связываться (реагировать) с малыми молекулами (эффекторами). В случае индуцированных ферментных систем репрессор связывается с оператором и блокирует транскрипцию олерона. Присутствие эффектора (индуктора) связывает (инактивирует) репрессор, и это приводит к тому, что происходит транскрипция и трансляция генов оперона. Другими словами, репрессор, соединенный с эффектором, теряет родство к оператору и не связывается с ним, а это сопровождается активацией оперона. В случае репрессибельных ферментов репрессор сам по себе является неактивным, т. е. не имеет родства к оператору и не блокирует транскрипцию оперона. Он активируется лишь в результате комбинации (соединения) с конечным продуктом в биосинтезе, в результате чего блокирует транскрипцию оперона. Следовательно, транскрипция оперона происходит в отсутствие эффектора (конечного продукта), тогда как присутствие эффектора сопровождается ингибированием оперона.
Регуляторные механизмы в случае индуцибельных и репрессибельных систем имеют негативный характер, т. к. подавляют синтез специфических белков. Регуляторный механизм оперирует на генетическом уровне, контролируя частоту синтеза мРНК. Опероны классифицируют на катаболизирующие (индуцибельные) и синтезирующие (репрессибельные).
Примером катаболизирующих оперонов является лактозный оперон (рис. 145), в состав которого входят структурные гены z, у и a, кодирующие b-галактозидазу, b-пермеазу и b-трансацетилазу, соответственно, ген-регулятор lac 1, кодирующий синтез репрессоpa (белок мм. 37 200, состоящий из четырех идентичных единиц, содержащих по 377 аминокислотных остатков), промоторы P2 и Р1, к которым присоединяется РНК-полимераза, и ген-оператор (О), управляющий функционированием структурных генов.

Механизм регуляции лактозного оперона заключается в том, что в отсутствие индуктора (лактозы) репрессор активен, т. е. продукт гена lac 1 связан с оператором O1 в соединении с двумя другими операторами (О2 и О3) и это блокирует транскрипцию полигенной мРНК, т. е. предупреждает движение РНК-полимеразы с промотора. Операторы О2 и О3, которых часто называют псевдооператорами, повышают связывание репрессора с оператором О1. Промотор P1 частично перекрывается вторым промотором P2, который в нормальных условиях не имеет существенного значения. РНК-полимераза может подходить к обоим промоторам, но основным все же является промотор Р1, поскольку движение РНК-полимеразы к промотору P2 ингибируется белком CRP, который при этом стимулирует транскрипцию с промотора P1. При наличии в среде индуктора репрессор теряет активность, т. е. связывается с индуктором, вследствие чего оператор становится свободным (деблокируется), и в результате движения РНК-полимеразы происходит транскрипция мРНК. При наличии в среде лактозы синтез ферментов увеличивается в 1000 раз. Инициация транскрипций требует наличия циклического АМФ и белка сгр. Следовательно, лактозный оперон контролируется негативно путем контроля частоты синтеза мРНК (рис. 146).
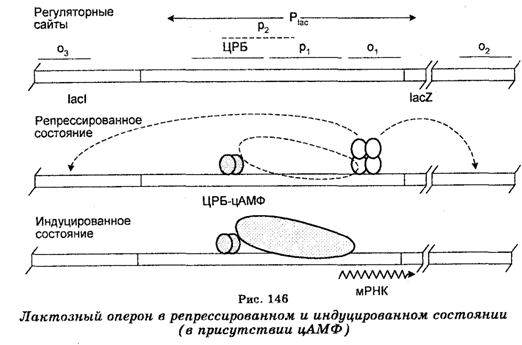
Однако возможна и позитивная регуляция лактозного оперона. Если вслед за индуктором в среду, где выращены бактерии,прибавить глюкозу, то наступает катаболитная репрессия (3-галак-тозидазы, сопровождающаяся снижением уровня цАМФ. Если же в культуру прибавить экзогенный цАМФ, то катаболитическая репрессия снимается и лактозный оперон будет функционировать нормально. Таким образом, цАМФ является позитивным регулятором (регуляция происходит в присутствии индуктора).
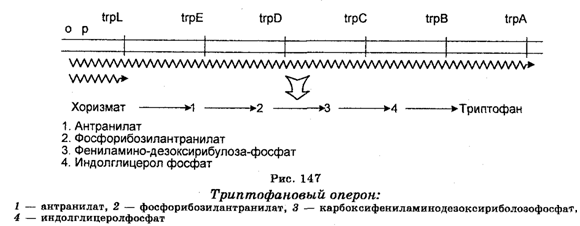
Примером биосинтезирующего оперона является триптофановый оперон, который состоит из 5 генов trpE, D, С, В и А, кодирующих ферменты, участвующие в биосинтезе триптофана (рис. 147). Первая химическая реакция, характерная для синтеза триптофана (как и для тирозина и фенилаланина), заключается в конденсации (накоплении) эритрозы-4-Ф и фосфоенолпирувата, необходимых для образования З-дезокси-арабино-гептулозоната-7-Ф (ДДГФ), катализируемого ДАГФ-синтетазой. Образующийся продукт (хоризмат) с помощью ферментов превращается в триптофан. Экспрессия этого оперона находится под негативным контролем репрессора, являющегося продуктом гена trp R. Репрессия конечным продуктом (трип-тофаном) заключается в том, что неактивный в свободном состоянии репрессор не мешает переходу РНК-полимеразы к промотору, т. е. не мешает транскрипции мРНК. Репрессор становится активным в результате связывания с конечным продуктом (триптофаном). Когда комплекс репрессор + триптофан связывает оператор, то это предупреждает преход РНК-полимеразы к промоторам, т. е. подавляет синтез мРНК. Более тонкий механизм заключается в том, что продукт гена trp R (репрессор) ингибирует синтез одной из ДАГФ-синтетаз, в частности, синтетазу АгоН, путем ингибирования ее мРНК, а следовательно, ингибирует и мРНК всего гистидинового оперона.
То, что существуют различные пути регуляции генов, подтверждается данными о регуляции оперона ara BAD, который ответственен за метаболизм арабинозы, встречающейся обычно в стенках клеток растений, а у микроорганизмов — являющейся одним из источников углерода и энергии. Арабиноза попадает в организм животных или человека с растительной пищей, где она метаболи-зируется микроорганизмами. Однако прежде она должна поступить в бактерии. Это поступление арабинозы в бактериальные клетки, т. е. транспорт арабинозы обеспечивается двумя независимыми системами, представляющими собой продукты генов ara Е и ara FG (рис. 148).
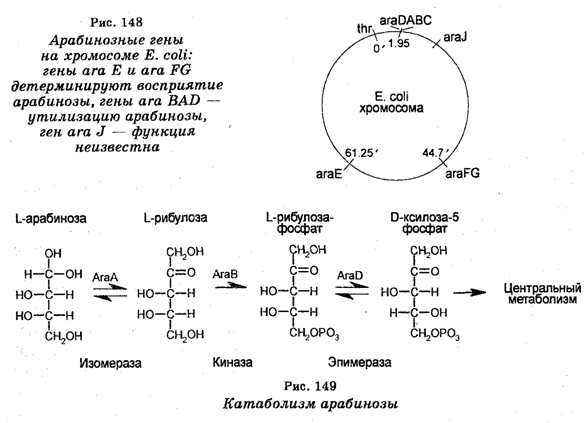
Система ara Е обладает низким родством к арабинозе, поэтому она используется лишь при очень высоких концентрациях этого углевода, тогда как система ara FG, обладающая высоким родством к арабинозе, используется при очень низких концентрациях углевода (порядка 10-7 М).
Гены, кодирующие ферменты, которые необходимы для катаболизма арабинозы, показаны на рис. 149. Катаболизм арабинозы начинается с того, что она вначале конвертируется синтезируемая под контролем гена ara А арабинозной изомеразой в L-рибулозу. Затем рибулоза фосфорилируется рибулозокиназой, являющейся продуктом гена ara В в L-рибулозу-б-фосфат. В свою очередь рибулозо-фосфат конвертируется в П-ксилозу-б-фосфат с помощью рибулозо-фосфатэпимеразы, являющейся продуктом гена ara D. Важно отметить, что гены транспортной системы, а также дополнительный арабинозоиндуцируемый ген и гены метаболизма арабинозы локализованы в разных районах хромосомной карты Е. coil. Все эти наборы генов регулируются позитивно арабинозой.

Обсуждение Биология с основами Экологии
Комментарии, рецензии и отзывы