Страница 70
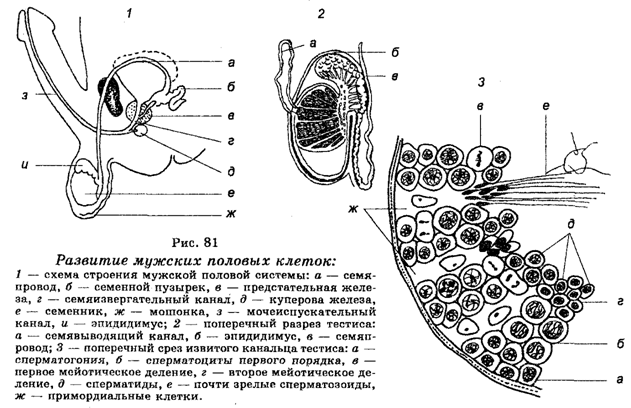
В период половой зрелости индивида часть сперматогоний перемещается во внутренний слой канальцев, где в результате мейоза они развиваются в клетки, получившие название сперматоцитов первого порядка (сперматоцитов I), затем в спер-матоциты второго порядка (сперматоциты II) и, наконец, в сперматиды, являющиеся гаплоидными зародышевыми клетками, дифференцирующимися в конечном итоге в зрелые сперматозоиды. Таким образом, в обобщенном виде можно сказать, что сперматогенез инициируется в диплоид-ных соматических клетках (сперматогони-ях), после чего следует период созревания зародышевых клеток, в котором происходит два деления ядер путем мейоза, приводящего к образованию сперматид.
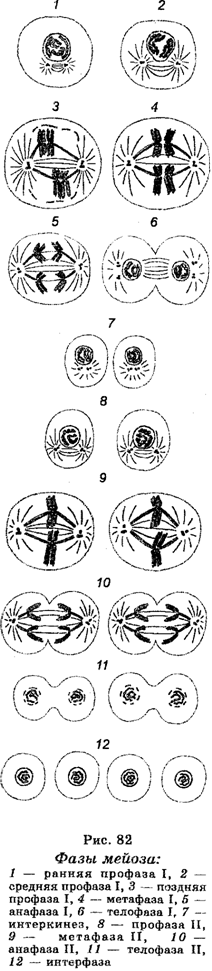 Мейоз в сперматогенезе протекает в несколько стадий (фаз). Между делениями имеются две интерфазы. Таким образом, мейотическое деление можно представить в виде серии следующих одно за другим событий, а именно: интерфаза I ® первое мейотическое деление (ранняя профаза I, поздняя профаза I, метафаза I, анафаза I, телофаза I) ® интерфаза II (интерокинез) ® второе мейотическое деление (профаза II, метафаза II, анафаза II, телофаза II). Процесс мейоза очень динамичен, поэтому микроскопические различия между разными стадиями отражают скорее не характер самих стадий, а скорее свойства хромосом на разных стадиях (рис. 82). Интерфаза I характеризуется тем, что в ней происходит репликация хромосом (удвоение ДНК), которая к началу ранней профазы I почти полностью завершается.
Мейоз в сперматогенезе протекает в несколько стадий (фаз). Между делениями имеются две интерфазы. Таким образом, мейотическое деление можно представить в виде серии следующих одно за другим событий, а именно: интерфаза I ® первое мейотическое деление (ранняя профаза I, поздняя профаза I, метафаза I, анафаза I, телофаза I) ® интерфаза II (интерокинез) ® второе мейотическое деление (профаза II, метафаза II, анафаза II, телофаза II). Процесс мейоза очень динамичен, поэтому микроскопические различия между разными стадиями отражают скорее не характер самих стадий, а скорее свойства хромосом на разных стадиях (рис. 82). Интерфаза I характеризуется тем, что в ней происходит репликация хромосом (удвоение ДНК), которая к началу ранней профазы I почти полностью завершается.
Первое мейотическое деление начинается в первичном сперматоците и характеризуется длинной профазой, которая состоит из переходящих одна в другую профазы I и профазы II. В профазе I различают пять главных стадий — лептонему, зигонему, пахинему, диплонему и диакинез.
На стадии лептонемы хромосомы в ядре представлены в виде тонких спирализованных нитей, содержащих многочисленные темноок-рашенные гранулы (хромомеры). Расщепления хромомер и нитей не отмечают, но считают, что хромосомы на уровне этой стадии являются двойными, т. е. диплоидными. Гомологи каждой хромосомной пары объединяются хромомерами вдоль их длины по принципу застежки.
На стадии зигонемы характерно установление синапсов между гомологичными хромосомами, в результате чего образуются спаренные хромосомы (биваленты). Хромосомы Х и Y ведут себя по сравнению с аутосомами несколько по-другому. Они конденсируются в темноокрашиваемые гетерохроматиновые тела, спариваемые в результате наличия гомологичных районов на их концах.
На стадии пахинемы, которая является наиболее долгой во времени стадией в мейотической профазе, происходит конденсация бивалентов и разделение каждой хроматиды надвое, в результате чего каждый бивалент представляет собой сложную спиральную структуру, состоящую из четырех сестринских хроматид (тетрад). В конце этой стадии начинается разделение спаренных хромосом-бивалентов. Теперь гомологичные хромосомы могут наблюдаться рядом. Поэтому в некоторых препаратах можно видеть четыре хромосомы, которые образуются в результате дупликации каждого гомолога, формирующего сестринские хроматиды. На этой стадии происходят обмены между гомологами и формирование хиазм.
На стадии диплонемы происходит укорачивание, утолщение и взаимное отталкивание сестринских хроматид, в результате чего хроматиды в биваленте почти разъединены. Разделение является неполным по той причине, что в каждой паре хромосом еще не расщеплена центромера. Что касается бивалентов, то они удерживаются на различных местах вдоль их длины с помощью хиазм, которые являются структурами, сформированными между гомологичными хроматидами в результате предыдущего кроссинговера между синаптически связанными гомологами. В хороших препаратах можно наблюдать от одной до нескольких хиазм в зависимости от длины бивалента. Каждая наблюдаемая на этой стадии хиазма представляет собой результат обмена, который встречался между несестринскими хроматидами в течение стадии пахинемы. Поскольку сжатие и отталкивание бивалентов усиливается, хиазмы двигаются к концам хромосом, т. е. происходит терминализа-ция хромосом. В конце диплонемы наступает деспирализация хромосом; гомологи продолжают отталкиваться друг от друга.
На стадии диакинеза, которая сходна с диплотеной, продолжается укорочение бивалентов и наступает ослабление (уменьшение) хиазм, вследствие чего формируются дискретные единицы в виде хроматид (четырех). Непосредственно после завершения этой стадии происходит растворение ядерной мембраны.
В метафазе I биваленты достигают наибольшей конденсации. Становясь овальными, они располагаются в экваториальной части ядра, где формируют экваториальные пластинки мейотической метафазы I. Форма каждого бивалента определяется числом и локализацией хиазм. У мужчин число хиазм на бивалент в метафазе I составляет обычно 1—5. Бивалент XY становится палочковидным в результате одиночной терминально расположенной хиазмы.
В анафазе I начинается движение противоположных центромер к противоположным полюсам клетки. В результате этого происходит разделение гомологичных хромосом. Каждая хромосома состоит теперь из двух хроматид, удерживаемых центромерой, которая не делится и остается интактной. Этим анафаза I мейоза отличается от анафазы митоза, при котором центромера подвергается разделению. Важно заметить, что благодаря кроссинговеру каждая хро-матида является генетически различной.
В стадии телофазы I хромосомы достигают полюсов, чем заканчивается первое мейотическое деление. После телофазы I наступает короткая интерфаза (интеркинез), в которой хромосомы деспи-рализуются и становятся диффузными, или телофаза I переходит прямо в профазу II второго мейотического деления. Ни в одном, ни в другом случае репликации ДНК не отмечается. После первого мейотического деления клетки называют спермат&цитами II порядка. Количество хромосом в каждой такой клетке снижается от 2п до п, но содержание ДНК еще не изменяется.
Второе мейотическое деление осуществляется в течение нескольких фаз (профаза II, метафаза II, анафаза II, телофаза II) и сходно с митотическим делением. В профазе II хромосомы вторичных спер-матоцитов остаются у полюсов. В метафазе П центромера каждой из двойных хромосом делится, обеспечивая каждую новую хромосому собственной центромерой. В анафазе II начинается формирование веретена, к полюсу которого двигаются новые хромосомы. В телофазе II второе мейотическое деление заканчивается, в результате чего каждый сперматоцит II порядка дает два спермати-да, из которых дифференцируются затем сперматозоиды. Как и во вторичном сперматоците, число хромосом в сперматиде является гаплоидным (п). Однако хромосомы сперматид являются одиночными, тогда как хромосомы вторичных сперматоцитов II являются двойными, будучи построенными из двух хроматид. Следовательно, ядро каждого сперматида имеет одиночный набор негомологичных хромосом. Вторичное мейотическое деление является делением митотического типа (экваториальным делением). Оно разделяет двойные сестринские хроматиды и отличается от редукционного деления, в котором гомологичные хромосомы разделены. Единственное существенное отличие от классического митоза заключается в том, что здесь имеется гаплоидный набор хромосом.

Обсуждение Биология с основами Экологии
Комментарии, рецензии и отзывы