2. факторы, влияющие на изменение частот аллелей в популяции
2. факторы, влияющие на изменение частот аллелей в популяции
Как уже говорилось, закон Харди-Вайнберга описывает популяции в состоянии покоя. В этом смысле он аналогичен первому закону Ньютона в механике, согласно которому любое тело сохраняет состояние покоя или равномерного прямолинейного движения, пока действующие на него силы не изменят это состояние.
Закон Харди-Вайнберга гласит: при отсутствии возмущающих процессов частоты генов в популяции не изменяются. Однако в реальной жизни гены постоянно находятся под воздействием процессов, изменяющих их частоты. Без таких процессов эволюция просто не происходила бы. Именно в этом смысле закон Харди-Вайнберга аналогичен первому закону Ньютона он задает точку отсчета, по отношению к которой анализируются изменения, вызванные эволюционными процессами. К последним относятся мутации, миграции и дрейф генов.
МУТАЦИИ КАК ИСТОЧНИК ГЕНЕТИЧЕСКОЙ ИЗМЕНЧИВОСТИ
Мутации служат основным источником генетической изменчивости, но их частота крайне низка. Мутирование процесс чрезвычайно медленный, поэтому если мутирование происходило бы само по себе, а не в контексте действия других популяционных факторов (например, дрейфа генов или миграции), то эволюция протекала бы невообразимо медленно. Приведем пример.
Предположим, существуют два аллеля одного локуса (т.е. два варианта одного гена) а и a. Допустим, что в результате мутации а превращается в a, а частота этого явления u на одну гамету за одно поколение. Допустим также, что в начальный момент времени (до начала процесса мутации) частота аллеля a равнялась р0. Соответственно, в следующем поколении u аллелей типа а превратятся в аллели типа a, а частота аллеля а будет равна р1 = р0 up0 = р0(1 u). Во втором поколении доля u оставшихся аллелей а (частота встречаемости которых в популяции теперь составляет р1) снова мутирует в a, а частота а будет равна р2= р1(1 u)= р0(1 u)´ (1 u)= р0(1 u)2. По прошествии t поколений частота аллеля а будет равна р0(1 u)t.
Поскольку величина (1 u). < 1, то очевидно, что с течением времени частота встречаемости аллеля а уменьшается. Если этот процесс продолжается бесконечно долго, то она стремится к нулю. Интуитивно эта закономерность достаточно прозрачна: если в каждом поколении какая-то часть аллелей а превращается в аллели а, то рано или поздно от аллелей типа а ничего не останется они все превратятся в аллели a.
Однако вопрос о том, как скоро это произойдет, остается открытым все определяется величиной u. В естественных условиях она чрезвычайно мала и составляет примерно 10-5. В таком темпе, для того чтобы изменить частоту аллеля а от 1 до 0,99, потребуется примерно 1000 поколений; для того чтобы изменить его частоту от 0,50 до 0,49 2000 поколений, а от 0,10 до 0,09 10 000 поколений. Вообще, чем меньше исходная частота аллеля, тем больше времени требуется на ее снижение. (Переведем поколения в годы: принято считать, что у человека смена поколений происходит каждые 25 лет.)
Разбирая этот пример, мы сделали предположение о том, что процесс мутирования односторонен а превращается в а, но обратного движения (a в а) не происходит. На самом деле мутации бывают как односторонние (а®a) , так и двухсторонние (а® a и a®а), при этом мутации типа а ®a называются прямыми, а мутации типа a® a называются обратными. Это обстоятельство, конечно же, несколько осложняет подсчет частот встречаемости аллелей в популяции.
Отметим, что частоты аллелей в естественных популяциях обычно не находятся в состоянии равновесия между прямыми и обратными мутациями. В частности, естественный отбор может благоприятствовать одному аллелю в ущерб другому, и в этом случае частоты аллелей определяются взаимодействием между мутациями и отбором. Кроме того, при наличии двухстороннего мутационного процесса (прямых и обратных мутаций) изменение частот аллелей происходит медленнее, чем в случае, когда мутации частично компенсируют снижение частоты встречаемости исходного дикого аллеля (аллеля а). Это еще раз подтверждает сказанное выше: для того чтобы мутации сами по себе привели к сколько-нибудь значительному изменению частот аллелей, требуется чрезвычайно много времени.
миграция
Миграцией называется процесс перемещения особей из одной популяции в другую и последующее скрещивание представителей этих двух популяций. Миграция обеспечивает «поток генов», т.е. изменение генетического состава популяции, обусловленное поступлением новых генов. Миграция не влияет на частоту аллелей у вида в целом, однако в локальных популяциях поток генов может существенно изменить относительные частоты аллелей при условии, что у «старожилов» и «мигрантов» исходные частоты аллелей различны.
В качестве примера рассмотрим некоторую локальную популяцию А, членов которой будем называть старожилами, и популяцию Б, членов которой назовем мигрантами. Допустим, что доля последних в популяции равна ц, так что в следующем поколении потомство получает от старожилов долю генов, равную (1 m), а от мигрантов долю, равную m. Сделаем еще одно допущение, предположив, что в популяции, из которой происходит миграция, средняя частота аллеля а составляет P, а в локальной популяции, принимающей мигрантов, его исходная частота равна р0. Частота встречаемости аллеля а в следующем (смешанном) поколении в локальной популяции (популяции-реципиенте) составит:
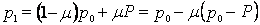 .
.
Другими словами, новая частота аллеля равна исходной частоте аллеля (р0), умноженной на долю старожилов (1 m) плюс доля пришельцев (m) , умноженная на частоту их аллеля (Р). Применив элементарные алгебраические приемы и перегруппировав члены уравнения, находим, что новая частота аллеля равна исходной частоте (р0) минус доля пришельцев М(m), умноженная на разность частот аллелей у старожилов и пришельцев (р0Р).
За одно поколение частота аллеля а изменяется на величину DР, Рассчитываемую по формуле: DР =р1 – р0 Подставив в это уравнение полученное выше значение р1 получим: 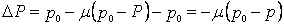 . Говоря иначе, чем больше доля пришельцев в популяции и чем больше различия в частотах аллеля а у представителей популяции, в которую иммигрируют особи, и популяции, из которой они эмигрируют, тем выше скорость изменения частоты этого аллеля. Отметим, что DР=0 только тогда, когда нулю равны либо m, т.е. миграция отсутствует, либо (р0Р), т.е. частоты аллеля а совпадают в обеих популяциях. Следовательно, если миграция не останавливается и популяции продолжают смешиваться, то частота аллеля в популяции-реципиенте будет изменяться до тех пор, пока р0 не будет равняться Р, т.е. пока частоты встречаемости а не станут одинаковыми в обеих популяциях.
. Говоря иначе, чем больше доля пришельцев в популяции и чем больше различия в частотах аллеля а у представителей популяции, в которую иммигрируют особи, и популяции, из которой они эмигрируют, тем выше скорость изменения частоты этого аллеля. Отметим, что DР=0 только тогда, когда нулю равны либо m, т.е. миграция отсутствует, либо (р0Р), т.е. частоты аллеля а совпадают в обеих популяциях. Следовательно, если миграция не останавливается и популяции продолжают смешиваться, то частота аллеля в популяции-реципиенте будет изменяться до тех пор, пока р0 не будет равняться Р, т.е. пока частоты встречаемости а не станут одинаковыми в обеих популяциях.
Как разница в частоте встречаемости аллеля в двух соседних популяциях изменяется во времени?
Допустим, что мы наблюдаем миграцию в течение двух поколений. Тогда после второго поколения различие в частотах встречаемости аллеля а в обеих популяциях будет равно
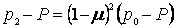 , (5.4)
, (5.4)
а после t поколений
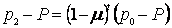
Эта формула чрезвычайно полезна. Во-первых, она позволяет рассчитать частоту аллеля а в локальной популяции (популяция старожилов) по прошествии t поколений миграции с известной скоростью m (при условии, что исследователю известны исходные частоты аллелей р0 и р1). И во-вторых, зная исходные частоты аллеля а в популяции, из которой мигрируют особи, и в популяции, в которую они мигрируют, конечные (послемиграционные) частоты аллеля а в популяции-реципиенте и продолжительность процесса миграции (t), можно рассчитать-интенсивность потока генов m.
Генетический след миграции. В США потомство от смешанных браков между белыми и черными принято относить к черному населению. Следовательно, смешанные браки можно рассматривать как поток генов из белой популяции в черную. Частота аллеля R°, контролирующего резус-фактор крови, составляет среди белых примерно Р= 0,028. В африканских популяциях, отдаленными потомками которых являются современные члены черной популяции США, частота этого аллеля равна р0 = 0,630. Предки современного черного населения США были вывезены из Африки примерно 300 лет назад (т.е. прошло примерно 10-12 поколений); для простоты примем, что t = 10. Частота аллеля R° современного черного населения США составляет pt = 0,446.
Переписав уравнение 5.5 в виде 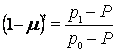 и подставив значения
и подставив значения
соответствующих величин, получим (1 m)'° = 0,694, m = 0,036. Таким образом, поток генов от белого населения США к черному шел со средней интенсивностью 3,6\% за одно поколение. В результате через 10 поколений доля генов африканских предков составляет примерно 60\% общего числа генов современного черного населения США и около 30\% генов (1 0,694 = 0,306) унаследовано от белых.
случайный дрейф генов
Любая природная популяция характеризуется тем, что она имеет конечное (ограниченное) число особей, входящих в ее состав. Этот факт проявляется в чисто случайных, статистических флуктуациях частот генов и генотипов в процессах образования выборки гамет, из которой формируется следующее поколение (поскольку не каждая особь в популяции производит потомство); объединения гамет в зиготы реализации «социальных» процессов (гибели носителей определенных генотипов в результате войн, бедствий, смертей до репродуктивного возраста); влияния мутационного и миграционного процессов и естественного отбора. Очевидно, что в больших популяциях влияние подобных процессов значительно слабее, чем в маленьких. Случайные, статистические флуктуации частот генов и генотипов называются популяционными волнами. Для обозначения роли случайных факторов в изменении частот генов в популяции С.Райт ввел понятие «дрейф генов» (случайный дрейф генов), а Н.П. Дубинин и Д.Д. Ромашов понятие «генетико-автоматические процессы». Мы будем использовать понятие «случайный дрейф генов».
Случайным дрейфом генов называется изменение частот аллелей в ряду поколений, являющееся результатом действия случайных причин, например, резким сокращением размера популяции в результате войны или голода. Предположим, что в некоторой популяции частоты двух аллелей а и ос равны 0,3 и 0,7 соответственно. Тогда в следующем поколении частота аллеля а может быть больше или меньше, чем 0,3, просто в результате того, что в наборе зигот, из которых формируется следующее поколение, его частота в силу каких-то причин оказалась отличной от ожидавшейся.
Общее правило случайных процессов таково: величина стандартного отклонения частот генов в популяции всегда находится в обратной зависимости от величины выборки чем больше выборка, тем меньше отклонение. В контексте генетики популяций это означает, что, чем меньше число скрещивающихся особей в популяции, тем больше вариативность частот аллелей в поколениях популяции. В небольших популяциях частота одного гена может случайно оказаться очень высокой. Так, в небольшом изоляте (дункеры в штате Пенсильвания, США, выходцы из Германии) частота генов групп крови АВО значительно выше, чем в исходной популяции в Германии [103]. И напротив, чем больше число особей, участвующих в создании следующего поколения, тем ближе теоретически ожидаемая частота аллелей (в родительском поколении) к частоте, наблюдаемой в следующем поколении (в поколении потомков).
Важным моментом является то, что численность популяции определяется не общим числом особей в популяции, а ее так называемой 'Эффективной численностью, которая определяется числом скрещивающихся особей, дающих начало следующему поколению. Именно эти особи (а не вся популяция в целом), становясь родителями, вносят генный вклад в следующее поколение.
Если популяция не слишком мала, то обусловленные дрейфом генов изменения частот аллелей, происходящие за одно поколение также относительно малы, однако, накопившись в ряду поколений, они могут стать весьма значительными. В том случае, если на частоты аллелей в данном локусе не оказывают влияния никакие другие процессы (мутации, миграции или отбор), эволюция, определяемая случайным дрейфом генов, в конечном счете приведет к фиксации одного из аллелей и уничтожению другого. В популяции, в которой действует только дрейф генов, вероятность того, что данный аллель будет фиксирован, равна исходной частоте его встречаемости. Иными словами, если аллель гена А в популяции встречается с частотой 0,1, то вероятность того, что в какой-то момент развития популяции этот аллель станет в ней единственной формой гена А, составляет 0,1. Соответственно, вероятность того, что в какой-то момент развития популяции зафиксируется аллель, встречающийся в ней с частотой 0,9, составляет 0,9. Однако для того, чтобы фиксация произошла, требуется достаточно много времени, поскольку среднее число поколений, необходимых для фиксации аллеля, примерно в 4 раза больше, чем число родителей в каждом поколении.
Предельный случай дрейфа генов представляет собой процесс возникновения новой популяции, происходящей всего от нескольких особей. Этот феномен известен под названием эффекта основателя (или «эффекта родоначальника»).
В. Маккьюсик описал эффект основателя у секты меннонитов (штат Пенсильвания, США). В середине 60-х этот популяционный изолят насчитывал 8000 чел., и почти все они произошли от трех супружеских пар, прибывших в Америку до 1770 г. Для них была характерна необычно высокая частота гена, вызывающего особую форму карликовости с полидактилией (наличием лишних пальцев). Это настолько редкая патология, что к моменту выхода книги Маккьюсика во всей медицинской литературе было описано не более 50 подобных случаев; в изоляте же меннонитов было обнаружено 55 случаев данной аномалии. Очевидно, случайно сложилось так, что один из носителей этого редкого гена и стал «основателем» повышенной его частоты у меннонитов. Но в тех их группах, которые живут в других районах США и ведут свое начало от других предков, эта аномалия не обнаружена [103].
Случайное изменение частот аллелей, являющихся разновидностью случайного дрейфа генов, феномен, возникающий в случае, если популяция в процессе эволюции проходит сквозь «бутылочное горлышко». Когда климатические или какие-то другие условия существования популяции становятся неблагоприятными, ее численность резко сокращается и возникает опасность ее полного исчезновения. Если же ситуация изменяется в благоприятную сторону, то популяция восстанавливает свою численность, однако в результате дрейфа генов в момент прохождения через «бутылочное горлышко» в ней существенно изменяются частоты аллелей, и затем эти изменения сохраняются на протяжении последующих поколений. Так, на первых ступенях эволюционного развития человека многие племена неоднократно оказывались на грани полного вымирания. Одни из них исчезали, а другие, пройдя стадию резкого сокращения численности, разрастались иногда за счет мигрантов из других племен, а иногда благодаря увеличению рождаемости. Наблюдаемые в современном мире различия частот встречаемости одних и тех же аллелей в разных популяциях могут в определенной степени объясняться влиянием разных вариантов процесса генетического дрейфа.
ЕСТЕСТВЕННЫЙ ОТБОР
Естественным отбором называется процесс дифференциального воспроизводства потомства генетически различными организмами в популяции. Фактически это означает, что носители определенных генетических вариантов (т.е. определенных генотипов) имеют больше шансов выжить и оставить потомство, чем носители других вариантов (генотипов). Дифференциальное воспроизводство может быть связано с действием разных факторов, среди которых называются смертность, плодовитость, оплодотворяемость, успешность спаривания и продолжительность репродуктивного периода, выживаемость потомства (иногда ее называют жизнеспособностью).
Мерой способности особи к выживанию и размножению является приспособленность. Однако, поскольку размер популяции обычно ограничен особенностями среды, в которой она существует, эволюционная результативность особи определяется не абсолютной, а относительной приспособленностью, т.е. ее способностью к выживанию и размножению по сравнению с носителями других генотипов в данной популяции. В природе приспособленность генотипов не постоянна, а подвержена изменению. Тем не менее в математических моделях значение приспособленности принимается за константу, что помогает при разработке теорий популяционной генетики. Например, в одной из наиболее простых моделей предполагается, что приспособленность организма полностью определяется структурой его генотипа. Кроме того, при оценке приспособленности допускается, что все локусы осуществляют независимые вклады, т.е. каждый локус может анализироваться независимо от других.
Выделяются три основных типа мутаций: вредоносные, нейтральные и благоприятствующие. Большинство новых мутаций, возникающих в популяции, являются вредоносными, так как снижают приспособленность их носителей. Отбор обычно действует против таких мутантов, и через некоторое время они исчезают из популяции. Данный тип отбора называется отрицательным (стабилизирующим). Однако cуществуют мутации, появление которых не нарушает функционирование организма. Приспособленность таких мутантов может быть так же высока, как и приспособленность аллелей-немутантов (исходных аллелей) в популяции. Эти мутации являются нейтральными, и естественный отбор остается равнодушным к ним, не действуя против них (дизруптивный отбор). При действии дизруптивного отбора внутри популяции обычно возникает полиморфизм несколько отчетливо различающихся форм гена (см. гл. IV). Третий тип мутантов появляется крайне редко: такие мутации могут повысить приспособленность организма. В этом случае отбор может действовать так, что частота встречаемости аллелей-мутантов может повыситься. Данный тип отбора называется положительным (движущим) отбором.
ПОДСТАНОВКА ГЕНОВ
Предельным случаем эволюционирования популяции является полное исчезновение из нее исходных аллелей. Подстановкой генов (полной заменой одного аллеля на другой) называется процесс, в результате которого аллель-мутант вытесняет исходно доминировавший аллель «дикого типа». Иными словами, в результате действия различных популяционных процессов (например, мутационного процесса, случайного дрейфа генов, отбора) в популяции обнаруживаются только аллели-мутанты: аллель-мутант появляется в популяции в единственном числе в результате единичной мутации, а затем, после смены достаточного количества поколений, его частота достигает 100\%, т.е. он фиксируется в популяции. Время, требующееся аллелю для достижения 100\% частоты встречаемости, называется временем фиксации. Очевидно, что не все аллели-мутанты достигают 100\% встречаемости и фиксируются в популяции. Обычно бывает наоборот: большинство аллелей-мутантов в течение нескольких поколений элиминируется. Вероятность того, что данный аллель-мутант зафиксируется в популяции, обозначается величиной, называемой вероятностью фиксации.
Новые мутанты возникают в популяциях постоянно, при этом в качестве одного из сопровождающих мутацию процессов разворачивается процесс подстановки генов, в которых аллель А заменяется новым аллелем Б, а тот в свою очередь заменяется аллелем В и т.д. Динамика этого процесса описывается понятием «скорость процессов подстановки генов», отражающим количество подстановок и фиксаций в единицу времени.

Обсуждение Психогенетика
Комментарии, рецензии и отзывы