5.10.1. структурно-функциональная организация коры
5.10.1. структурно-функциональная организация коры
А. Кора головного мозга представляет собой многослойную нейронную ткань со множеством складок общей площадью в обоих полушариях примерно 2200 см2, что соответствует квадрату со сторонами 47 х 47 см; ее объем соответствует 40\% от массы головного мозга, толщина колеблется от 1,3 до 4,5 мм, а общий объем равен 600 см3. В состав коры головного мозга входит 109 –1010 нейронов и множество глиальных клеток, общее число которых пока не известно.
В коре выделяют 6 слоев (I-VI), каждый из которых состоит из пирамидных и звездчатых клеток (рис. 5.12). Главная особенность пирамидных клеток заключается в том, что их аксоны выходят из коры и оканчиваются в других корковых или иных структурах. Название звездчатых клеток также обусловлено их формой; их аксоны оканчиваются в коре, т.е. речь идет о корковых интернейронах.
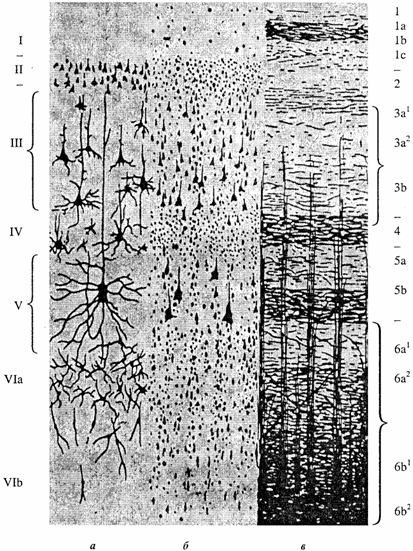
Рис. 5.12. Полусхематичное изображение слоев коры головного мозга (по К. Brodmann, Vogt; с изменениями): а основные типы нервных клеток (окраска по Гольджи); бтела нейронов (окраска по Нисслю); в общее расположение волокон (миелиновых оболочек). Слои пронумерованы от поверхности вглубь. Приведены две наиболее распространенные системы нумерации. (Объяснение в тексте.)
В I-IV слоях воспринимаются и обрабатываются поступающие в кору в виде импульсов сигналы. Покидающие кору эфферентные пути формируются преимущественно в V-VI слоях. Деление коры на различные поля проведено К.Бродманом (1909) на основе цитоархитектонических признаков формы и расположения нейронов. Многие из полей характеризуются функциональными и нейрохимическими особенностями. К.Бродман выделил 11 областей, включающих 52 поля: 1) постцентральную область (поля 1, 2, 3 и 43); 2) прецентральную область (поля 4 и 6); 3) лобную область (поля 8, 9, 10, 11, 12, 44, 45, 46 и 47); 4) островковую область (поля 13, 14, 15 и 16); 5) теменную область (поля 5, 7, 40 и 39); 6) височную область (поля 20, 21, 22, 36, 37, 38, 41, 42 и 52); 7) затылочную область (поля 17, 18 и 19); 8) поясную область (поля 23, 31, 24, 32, 33 и 25); 9) ретросплениальную область (поля 26, 29 и 30); 10) гиппокампову область (поля 27, 28, 34, 35 и 48); 11) обонятельную область (поле 51, nucleus amygdalae и обонятельный бугорок).
Более точно поля коры большого мозга описаны в трудах Московского научно-исследовательского института мозга, однако основная нумерация (52 поля) сохранена по Бродману (подробнее см. [9]).
Согласно гистологическим данным, элементарные нейронные цепи расположены перпендикулярно поверхности коры. Электрофизиологические исследования с погружением микроэлектродов перпендикулярно поверхности соматосенсорной коры показали, что все встречаемые на пути нейроны отвечают на раздражитель только одного качества (вида), например на свет. При погружении электрода под углом на его пути попадаются нейроны разной модальности. Это свидетельствует о том, что в коре мозга имеются функциональные объединения нейронов, расположенные в цилиндрике диаметром 0,5-1,0 мм. Эти объединения были названы нейронными колонками. Соседние нейронные колонки могут частично перекрываться и осуществлять саморегуляцию по типу возвратного торможения, а также взаимодействовать друг с другом по механизму латерального торможения.
Б. По своим биофизическим свойствам нейроны коры сходны с другими нервными клетками. Потенциал покоя пирамидных клеток составляет от -50 до -80 мВ, а амплитуда их потенциала действия -60-100 мВ при длительности 0,5-2 мс. Потенциалы действия в нейронах коры возникают в области аксонного холмика и распространяются не только по аксону, но также по телу и дендритам клетки (по крайней мере проксимальным). Поскольку вслед за потенциалом действия в этих нейронах не возникает заметной следовой гиперполяризации, частота их импульсации достигает 100 Гц. В нейронах коры даже у бодрствующих животных частота импульсации, вызванной постсинаптическими потенциалами, низкая, как правило, меньше 10 Гц, а нередко не достигает и 1 Гц. Пороговый потенциал (ΔV) клеток коры большого мозга колеблется и составляет 3-10 мВ, т.е. эти клетки обладают достаточно высокой возбудимостью.
В. Кортикализация функций возрастание в филогенезе роли коры большого мозга при анализе и регуляции функций организма и подчинение себе нижележащих отделов ЦНС. Выключение коры больших полушарий у обезьян и у человека приводит к потере не только локомоции, но и выпрямительных рефлексов, которые у других млекопитающих сохраняются.

Обсуждение Нейрофизиология и высшая нервная деятельность детей и подростков
Комментарии, рецензии и отзывы