Страница 117
Однако у животных ряда видов пресмыкающихся пол контролируется не генотипом, а средой. Например, у многих черепах пол детерминируется температурой инкубации отложенных яиц (женские особи развиваются при повышенных температурах инкубации, мужские — при пониженных). А у аллигаторов, крокодилов и ящериц отдельных видов мужской пол детерминируется повышенными температурами. Детерминирование пола средой у позвоночных является исключением из общего правила генетического контроля пола.
Еще в 1910 г. Т. Морган (1886-1945) установил, что половые хромосомы участвуют также в контроле и ряда признаков. В его экспериментах, заключающихся в скрещиваниях белоглазых дрозофил-самок с красноглазыми дрозофилами-самцами, оказалось, что гибридное потомство состояло из красноглазых самок и белоглазых самцов. Однако среди гибридов встречались отдельные красноглазые самцы и белоглазые самки, появление которых было неожиданным, исходя из особенностей передачи хромосомы Х и нахождения на ней локуса гена, контролирующего белый цвет глаз. Анализируя этот случай, К. Бриджис (1916) предположил, что «необычные» белоглазые самцы несут по две Х-хромосомы, унаследовав их от своих «матерей», тогда как «необычные» красноглазые самцы имеют одну Х-хромосому, унаследовав ее от «отцов». Объясняя это предположение, К. Бриджис допустил, что исходные белоглазые самки продуцировали «необычные» гаметы, содержащие либо две хромосомы XX, либо ни одной хромосомы Х (наряду с гаметами, обычно содержащими по одной хромосоме X) в результате нерасхождения при мейозе их хромосом XX. Если такие «необычные» гаметы подвергались оплодотворению сперматозоидами красноглазых самцов, то это приводило к образованию зигот четырех типов: а) белоглазых самок, б) красноглазых самцов, в) особей с 3 хромосомами Х нежизнеспособных, г) особей без хромосомы Х (также нежизнеспособных). Назвав это явление первичным нерасхождение хромомсом (рис. 128). К. Бриджис затем цитологически показал, что в соматических клетках «необычных» белоглазых самок F, вместо хромосом XX присутствуют две Х и одна Y-хромосома (XXY).
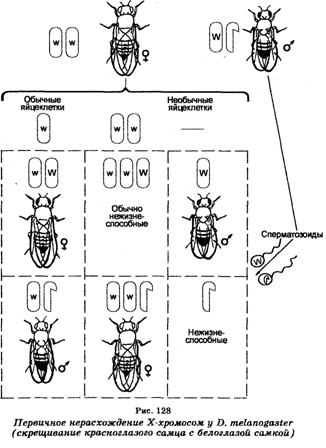
«Необычные» красноглазые самцы, не имеющие хромосомы Y, т. е. моносомики ХО, оказались в дальнейших экспериментах стерильными, тогда как белоглазые самки XXY были плодовитыми. Скрещивая последних с нормальными красноглазыми самцами и анализируя их потомство F1, К. Бриджис открыл вторичное нерасхождение хромосом при мейозе у самок XXY (рис. 129), поскольку среди организмов F, 96% самок имели красные глаза и 4% — белые, тогда как среди самцов 96% были белоглазыми и 4% — красноглазыми. В этом случае при мейозе самки XXY продуцируют гаметы четырех типов, в которых хромосомы Х и Y распределяются неодинаково (яйцеклетки с хромосомой X, яйцеклетки, имеющие по одной хромосоме Х и Y, яйцеклетка с двумя хромосомами Х и яйцеклетки лишь с хромосомами Y).
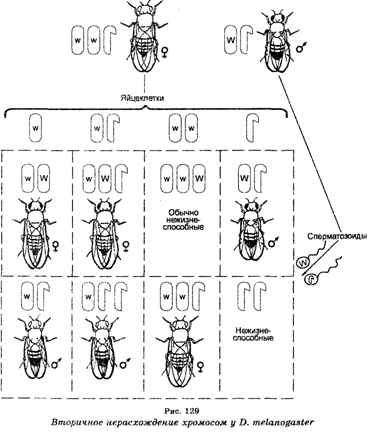
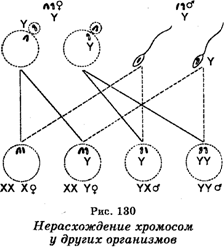
Оплодотворение яйцеклеток каждого из этих типов сперматозоидами нормального красноглазого самца приводило к образованию зигот восьми типов. Однако эти зиготы будут формироваться не с одинаковой частотой, поскольку мейоз у самок XXY связан с тем, что расхождение двух хромосом Х имеет место лишь в 92% овоцитов Объясняя вторичное нерасхождение хромосом, К. Бриджис допустил, что все белоглазые и отдельные красноглазые самки должны обладать не только парой хромосом X, но и хромосомой Y, тогда как красноглазые самцы в отличие от таких самцов, отмечаемых при первичном нерасхождении, должныиметь Y-хромосому и быть плодовитыми. Что касается отдельных белоглазых самцов, то они должны обладать одной хромосомой Х и двумя хромосомами Y. Эти допущения К. Бриджис доказал цитологически, найдя в соматических клетках дрозофилы хромосомы во всех сочетаниях, приведенных выше. Это открытие было показано на примере и других организмов (рис. 130).
 Открытие нерасхождения хромосом явилось окончательным доказательством в начале нашего века того, что гены локализованы на хромосомах. Однако теперь, когда с тех пор прошло много времени и когда к этому открытию возможен другой подход, можно сформулировать другое заключение. Фундаментальное значение открытия нерасхождения хромосом состоит в том, что оно положило начало изучению хромосомных мутаций и их механизмов. Можно добавить также, что оно означало объединение генетики и цитологии, т. е. формирование цитогенетики, оказавшейся в наше время исключительно плодотворной при изучении наследственности и наследственной патологии животных, растений и особенно человека (см. гл. XIII).
Открытие нерасхождения хромосом явилось окончательным доказательством в начале нашего века того, что гены локализованы на хромосомах. Однако теперь, когда с тех пор прошло много времени и когда к этому открытию возможен другой подход, можно сформулировать другое заключение. Фундаментальное значение открытия нерасхождения хромосом состоит в том, что оно положило начало изучению хромосомных мутаций и их механизмов. Можно добавить также, что оно означало объединение генетики и цитологии, т. е. формирование цитогенетики, оказавшейся в наше время исключительно плодотворной при изучении наследственности и наследственной патологии животных, растений и особенно человека (см. гл. XIII).
§55 Сцепление и кроссинговер
Рассматривая 2-й закон наследственности, мы видели, что перераспределение генов имеет независимый характер и связано с прохождением отцовского и материнского членов каждой пары хромосом при мейозе в разные гаметы. Следовательно, в независимое перераспределение или рекомбинацию генов вовлекаются те наборы генов, которые располагаются на разных хромосомах. Такую рекомбинацию называют свободной рекомбинацией. Однако еще в 1910 г. Т. Морган установил, что гены, располагающиеся на одной хромосоме, сцеплены между собой, причем степень сцепления генов зависит от расстояния между ними. Следовательно, Т. Морган установил, что 2-му закону наследственности подчиняются лишь гены, локализованные на разных хромосомах. Однако значение этих наблюдений заключалось также и в том, что они послужили основой для формулирования в дальнейшем Т. Морганом и его сотрудниками представлений о линейном расположении генов на хромосомах. На основе этих представлений стали создавать генетические карты хромосом не только дрозофилы, но и других организмов.
Сцепление генов хорошо изучено в случаях многих видов животных и растений, показано, что оно присуще как женским, так и мужским особям. Чтобы лучше понять природу сцепления, рассмотрим его на описанном шведским генетиком Хатчисоном примере сцепления генов, детерминирующего форму и окраску семян кукурузы.
Обозначим символом С доминантный аллель, детерминирующий окраску семян, символом с — рецессивный аллель, детерминирующий отсутствие окраски семян, символом S — доминантный аллель, детерминирующий нормальную форму семян и символом s — рецессивный аллель, детерминирующий морщинистость семян (рис. 131). Если скрещивать растения CS/CS и cs/cs, то гибриды F,, обладая генотипом CS/cs, будут давать окрашенные семена нормальной формы. В соответствии со вторым законом наследственности при независимом перераспределении генов С и S гибриды F1 должны были продуцировать гаметы CS, Cs, cS и cs, причем в равных количествах, оплодотворение которых гаметами организмов cess (скрещивание гибридов F1 с исходным родителем cess) должно было дать потомство четырех фенотипических типов в отношении 1:1:1:1. Между тем скрещивание гибридов F1 с исходными родительскими организмами cess приводило к получению организмов в совершенно иных количественных отношениях, а именно:

Обсуждение Биология с основами Экологии
Комментарии, рецензии и отзывы