Страница 128
тРНК характеризуются исключительно высокой специфичностью, что проявляется в их антикодоновых последовательностях, соответствующих кодонам, доступности для распознания нужной аминоацил-тРНК-синтетазой и в точности связывания с сайтами А и Р на рибосомах.
Инициация, элонгация и терминация полипептидного синтеза находятся под генетическим контролем.
Наряду с кодонами, детерминирующими последовательность аминокислот, существуют кодоны, определяющие начало и конец чтения иРНК. В синтезе белка существенная роль принадлежит N-концевой аминокислоте формилметионину и его тРНК. N-формилметионил-тРНК (ОНС×NH×CH×(CH2×CH2× SCH3)CO×О×тРНК) образуется в результате формилирования а-ами-ногруппы метионина NH2×СН(СН2×СН2×SCH3)×СО×ОН в метионил-тРНК. Поскольку формулирование характерно только для метионина и катализируется ферментом трансформилазой, то считают, что формилметионин-тРНК является инициатором синтеза полипепти-да. Это означает, что все полипептиды в процессе синтеза начинаются с метионина. N-формилметионин является N-концевой аминокислотой всех белков.
Инициация полипептидной цепи у кишечной палочки начинается с образования комплекса между мРНК, формилметионин-тРНК и рибосомной единицей 30 8, которое обеспечивается факторами (белками) инициации IF1, IF2 и IF3, а также ГТФ. Этот комплекс вступает в комбинацию с 50 S-рибосомной единицей, в результате чего формилметионин-тРНК становится связанной с пептидиловым сайтом. Энергия для этого обеспечивается гидролизом одной молекулы ГТФ. Кодоны АУГ, ГУА и ГУГ на 5'-конце или рядом с ним направляют включение N-формилметионина в качестве N-концевой аминокислоты белка. Можно сказать, что эти кодоны являются специфическими инициаторами белкового синтеза. Наиболее активным является кодон АУГ.
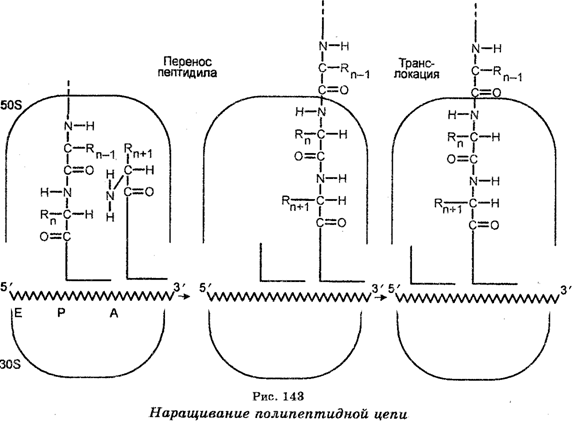
Элонгация (удлинение) полипептидной цепи обеспечивается белковыми факторами элонгации ef-TS и EF-Tu, а также гидролизом одной молекулы ГТФ, а движение молекулы мРНК с одного сайта рибосомы на другой обеспечивается фактором элонгации EF-G и гидролизом одной молекулы ГТФ. Каждый раз мРНК движется на три нуклеотида. У бактерий частота элонгации составляет 16 аминокислот в секунду. Это означает, что рибосомы двигаются вдоль мРНК со скоростью 48 нуклеотидов в секунду.
Терминация (окончание) синтеза детерминируется стоп-кодо-нами УАГ, УАА и У ГА. Когда один из этих кодонов подойдет к А-сайту рибосомы, то полипептид, тРНК в Р-сайте и мРНК освобождаются, а рибосомные субъединицы диссоциируют. Окончание синтеза белка связано с активностью белковых факторов освобождения — RF-1 и RF-2. Диссоциировав, рибосомные субъединицы начинают трансляцию другой молекулы мРНК. Большинство мРНК симультанно транслируется несколькими рибосомами (полисомами). Например, цепь гемоглобина из 150 аминокислот синтезируется на пентарибосомном комплексе. У прокариот синтез и трансляция мРНК происходят в направлении от 5'-конца к 3'-концу. Далее, у них нет ядерной мембраны. Поэтому трансляция мРНК начинается еще до завершения ее синтеза. Напротив, у эукариот транскрипция и трансляция разделены во времени, поскольку требуется время для перехода мРНК из ядра через ядерную мембрану в цитоплазму.
Синтез белков является исключительно точным механизмом. Обобщенные результаты исследований частоты ошибок в белковом синтезе показывают, что одна ошибка, т. еодно включение «неправильной» аминокислоты, происходит лишь на каждые 10 000 включенных аминокислот. Точность механизма белкового синтеза обеспечивается точностью связывания аминокислот со своими тРНК и точностью спаривания кодонов мРНК с антикодонами тРНК.
§ 61 Митохондриальный и хлоропластный
генетические коды
Помимо генетического кода, который содержится в ядерной ДНК, существует генетический код, содержащийся в ДНК мито-хондрий животных и человека, а также в ДНК хлоропластов растений. В митохондриях и хлоропластах помимо ДНК существуют и другие структуры, которые в совокупности с ДНК образуют самостоятельный аппарат синтеза белков. Размеры митохондриальных рибосом очень варьируют. В частности, размеры митохондриальных рибосом человека составляют 60 S.
Для митохондриального генетического кода характерны те же структуры и свойства и те же механизмы транскрипции и трансляции, что и в случае ядерного генетического кода. Однако известны и отличия. В митохондриальной ДНК все нуклеотиды входят в состав кодонов, кодирующих либо белки, либо рРНК и тРНК. Для трансляции используется только 22 тРНК (в отличие от 31 тРНК в ядерном коде и 30 тРНК в хлоропластном коде), причем отдельные молекулы тРНК могут узнавать любое основание, находящееся в кодоне в третьем положении. Митохондриальная ДНК человека и других млекопитающих содержит 64 кодона, из которых 4 являются стоп-кодонами.
Известно содержание антикодонов всех 22 тРНК (табл. 21). Каждый антикодон в случае митохондриального генетического кода способен спариваться с несколькими кодонами мРНК. Например, антикодон УАГ спаривается с кодонами ЦУУ, ЦУЦ, ЦУА и ЦУТ, кодирующими лейцин. 22 антикодона тРНК спариваются с 60 кодонами иРНК. Установлено, что митохондриальные тРНК подвержены «редактированию» (модификации транспорта тРНК) путем полиаденилирования, в результате чего создаются антикодоны терминации.
Таблица 21
Антикодоны в митохондриальном генетическом
коде млекопитающих*
ГАА
Фенилаланин
ГУА
тирозин
УАА
Лейцин
ГУГ
гистидин
УАГ
Лейцин
УУГ

Обсуждение Биология с основами Экологии
Комментарии, рецензии и отзывы